Bab 11. Avian Influenza berpatogenitas rendah (LPAI)
Oleh Daniel R. Perez dan J. J. (Sjaak) de Wit
Diterjemahkan oleh: Agna D. Lantria
DISCLAIMER
Artikel terjemahan ini dimaksudkan bagi pembaca berbahasa Indonesia untuk dapat mempelajari artikel keilmuan terkait. Penerjemah tidak mendapatkan keuntungan apapun dari kegiatan penerjemahan ini.
Famili Orthomyxoviridae meliputi enam genera, yaitu virus Influenza A, virus Influenza B, virus Influenza C, Isavirus, Thogotovirus, dan Quaranjavirus [181]. Hanya virus-virus dari genus virus Influenza A yang diketahui dapat menginfeksi burung-burung [6]. Burung-burung air (termasuk kedalam ordo Anseriformes dan Charadriiformes) dianggap sebagai hospes alami dan reservoir virus influenza A (IAV), sementara burung-burung galinaseus (unggas) tidak dianggap sebagai hospes alami IAV [6]. Berdasarkan perbedaan antigenic pada dua glikoprotein permukaan utama, yaitu hemagglutinin (HA) dan neuraminidase (NA), IAV kemudian dibagi menjadi subtipe-subtipe. Sampai saat ini, IAV yang berasal dari avian yangt termasuk kedalam 16 subtipe HA (H1-H16) dan 9 subtipe NA (N1-N9) telah diisolasi dari burung [6]. Meskipun Batasan hospes yang mencegah virus IAV menyebrang dari hospes alami kepada unggas mungkin saja lebih rendah daripada batasan yang mencegah virus-virus ini berpindah ke spesies hewan lainnya, persebaran dan perpetuasi virus-virus ini pada unggas diperkirakan akan membawa pada perubahan adaptif yang mempengaruhi virulensi, transmisi, dan kisaran hospes. Dari sudut pandang manajemen penyakit dan pelaporan, istilah “avian influenza” didefinisikan oleh Organisasi Kesehatan Hewan Dunia (OIE) Terrestrial Animal Health Code sebagai infeksi pada unggas yang disebabkan oleh setiap virus avian influenza berpatogenisitas tinggi (HPAI), dan oleh subtipe H5 dan H7 dengan patogenitas rendah (LPAI H5/H7), sedangkan infeksi yang disebabkan oleh subtipe-subtipe lainya (H1-H4, H6, dan H8-H16) hanya disebut sebagai influenza A [203]. Untuk menyederhanakannya, di Bab ini istilah virus LPAI digunakan untuk merujuk kepada IAV avian yang memiliki patogeniras rendah tanpa memperhatikan subtipenya, dan yang memiliki indeks patogenitas intravena (intravenous pathogenicity index/ IVPI) kurang dari 1.2. Perlu juga diperhatikan bahwa bayak negara didunia tidak memiliki sumber daya yang cukup atau otoritas pengatur yang dapat melakukan investigasi dan melaporkan keberadaan dan prevalensi virus LPAI se;ain H5 atau H7. Karena itulah daftar yang ada kemungkinan tidaklah lengkap dan biar terhadap subtipe-subtipe yang wajib dilaporkan (notifiable), yaitu H5 dan H7. Pada Bab ini fokus utama adalah pada virus LPAI yang telah menyebabkan wabah pada unggas yang lolos dari upaya-upaya penanggulangan dan telah menjadi endemic dan/atau menjadi resiko kesehatan masyarakat yang signifikan. Utamanya kita akan mendiskusikan tentang virus LPAI H9N2 dan H7N9 di Asia dan H5N2 di Meksiko.
Burung-burung liar sebagai reservoir alami dan sumber masuknya virus kedalam sistem pertanian
Replikasi virus IAV pada unggas air liar dan burung-burung pesisir terjadi utamanya pada saluran pencernaan dan kadang pada saluran pernafasan [57, 240, 241, 291]. Itik dapat mengeluarkan virus selama 30 hari dan dapat mengekskresikan 108mean embryo infectious doses per 1 ml (EID50/Ml) feses [292], yang mengkontaminasi air permukaan, rawa-rawa dan dataran pesisir. Transmisi diduga terjadi secara efektif melalui rute fekal-oral melalui ekskresi virus di dalam air. Transmisi fekal-oral dan pergerakan burung-burung air liar dan burung-burung pesisir adalah mekanisme yang memungkinkan virus untuk dapat bertahan dan menyebar di alam [58, 240, 241]. Ordo-ordo burung lain mungkin saja juga terinfeksi, akan tetapi virus AI tampaknya tidak beredar secara alam dalam populasi mereka [290]. Akan tetapi, penting untuk dikenali bahwa beberapa burung-burung ini dapat bertindak s=baik sebagai vector biologis maupun mekanis untuk pergerakan virus AI antara burung air sebagai reservoir dan unggas domestik. Di Belanda, peternakan komersial ayam petelur yang memiliki kandang luar ruangan (outdoor) telah memperlihatkan meningkatnya resiko masuknya virus LPAI sebanyak lebih dari sepuluh kali lipat jika dibandingkan dengan peternakan layer dalam ruangan (indoor) [88]. Karena itulah, burung-burung air dan burung-burung pesisir sebagai reservoir beserta lingkungan yang menjadi tempat hidupnya, serta setiap benda mati ataupun benda hidup (termasuk manusia) yang berbagi tempat hidup dengan burung-burung ini dapat menjadi sumber virus AI terhadap unggas domestik.
Perpetuasi virus avian influeza dalam hospes yang bukan alamiahnya: peranan pasar unggas hidup (Live Bird Market/LBM)
Reservoir virus paling utama dari virus LPAI adalah pasar unggas hidup (live bird market/LBM)[24, 272]. Jika dikombinasikan dengan sistem produksi unggas pedesaan yang memuat beragam unggas yang diumbar, LBM memaikan peranan besar dalam kemunculan dan bertahan hidupnya virus LAPI (dan virus HPAI) [215]. Pada unggas, infeksi LPAI dapat mempengaruhi baik saluran pencernaan maupun saluran pernafasan, atau keduanya. Pada awalnya, transmisi virus PLAI bisa jadi tidak seefektif penularan pada unggas akuatik, namun akses bersama terhadap tempat pakan dan tempat minum, dimana terdapat kesempatan adanya kontaminasi dari baik feses maupun sekresi saluran nafas, memfasilitasi transmisi di dalam flok-flok unggas [91]. LBM yang memiliki kepadatan unggas tinggi dan memiliki aktivitas manusia yang sibuk menyediakan lingkungan yang ideal untuk virus LPAI untuk dapat beradaptasi secara penuh dan berkembang pesat pada unggas [5, 50, 137, 193, 238, 255, 272, 312].
Teka-Teki dalam sistem pertanian: sistem perikanan terintegrasi
Berpusat pada kolam-kolam ikan, kegiatan perikanan terintegrasi adalah sebuah sistem produksi ikan yang berpadu dengan kegiatan-kegiatan pertanian dan peternakan hewan. Perikanan terintegrasi kemungkinan adalag sistem pertanian paling ramah lingkungan di planet ini. prinsip perikanan terintegrasi melibatkan pemeliharaan ikan bersama hewan ternak dan atau komoditas pertanian. Dalam sistem ini, produk buangan dari satu sistem biologi bertindak sebagai sumber nutrisi untuk sistem biologi kedua. Dalam sistem terintegrasi yang semacam ini, subsistem-subsistemnya sangat terkait dalam hubungan yang saling menguntungkan dalam satu area terbatas, meminimalisir biaya-biaya produksi akan tetapi menghasilkan berbagai produk pertanian yang beragam, termasuk (meskipun tidak terbatas pada) ikan, daging, telur, sayur mayur, kayu untuk bahan bakar, dan makanan ternak. Terdapat kombinasi yang kompleks yang terdiri dalam beberapa sistem. Di Asia, sistem perikanan umumya mengintegrasikan unggas-unggas air maupun unggas darat, juga babi yang ditempatkan berdekatan satu sama lain dan berkontak dengan kehidupan liar, termasuk burung liar akuatik. Integrasi ini menciptakan sebuah teka teki besar, yaitu bagaimana caranya mempertahankan sistem pertanian yang efisien tanpa meningkatkan resiko masuknya pathogen-patogen baru ataupun yang bersifat zoonotic [230]. Meskipun telah dibantah melalui beberapa laporan, virus LPAI adalah lambing dari pathogen yang mampu mencari lingkungan yang ideal untuk beralih hospes [80, 8]. Pergerakan hewan hidup dan atau peralatan yang terkontaminasi masuk dan keluar sistem-sistem ini dan kedalam pasar-pasar hewan menambah pada kompleksitas yang memajukan kemunculan pathogen-patogen baru (novel) yang berpotensi zoonosis [238].
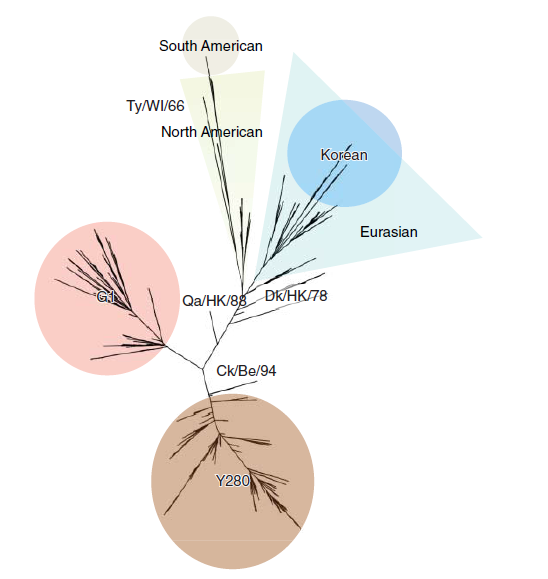
Mulai dari introduksi awal virus-virus H9N2 kepada unggas di China, tidak ada virus LPAI lain yang telah menunjukkan ketahanan yang sama. Penyebarannya telah mencapai spesies-spesies unggas domestik di berbagai negara di Asia, Timur Tengah, dan Afrika. Dua keturunan filogeografikal yang berbeda dari virus H9N2 telah dideskripsikan, yaitu keturunan Amerika dan Eurasia (Gambar 11.1). Dari dua keturunan utama ini selanjutnya klaster-klaster kemudian dapat diidentifikasi, meliputi klaster Amerika Selatan pada unggas liar [303]., dan dua klister utama virus-virus yang telah beradaptasi pada unggas di Asia [47].
Desktripsi awal virus LPAI dari subtipe H9N2 adalah sejak dari terjadinya wabah pada Kalkun di Wisconsin utara, Amerika Serikat pada Februari 1966 [100, 245]. Mortalitas yang rendah, batuk, bersin-bersin, dan berkurangnya produksi telur dan berkurangnya kemampuan menetasnya telur secara nyata menandai terjadinya wabah. Virus-virus H9 belum memiliki keturunan yang stabil pada unggas di Amerika Utara, meskipun sebanyak 16 wabah yang teridentifikasi berhubungan dengan subtipe H9 pada kalkun telah terdokumentasikan antara tahun 1981 sampai 1996 [90]. Detail wabah-wabah ini dan wabah-wabah LPAI lainnya pada unggas di Amerika Serikat, terutama pada wilayah-wilayah negara bagian penghasil utama kalkun yaitu Minnesota dan Wisconsin, telah dijelaskan pada buku terbitan edisi sebelumnya [89]. Kemunculan virus HPAI H5N1 dan infeksi pada manusia yang terjadi setelahnyadi tahun 1997 menyebabkan perubahan pada praktek-praktek peternakan umbaran di sebagian Amerika Serikat. Di Minnesota, produksi kalkun umbaran berhenti total mulai dari tahun 1998 dan seterusnya, tercatat sebesar 0.5% dari flok kalkun di negara bagian tersebut. Efek dari perubahan ini adalah berkurangnya wabah LPAI secara signifikan. Pada saat tulisan ini dibuat, pencegahan wabah HPAI sebagian besarnya dicapai melalui pencegahan paparan terhadap virus influenza dengan menghindari kontak langsung maupun tidak langsung dengan burung-burung air dan burung-burung pesisir dan lingkungan dimana burung tersebut tinggal. Hanya terdapat isolasi sporadis virus dari subtipe H9 dan beberapa virus LPAI lain ditemukan pada unggas di Amerika Serikat. Sebaliknya, virus-virus H9 seringkali ditemukan pada itik liar dan burung-burung pesisir di Amerika Utara [89].
Di Asia, virus-virus dari subtipe H9 (biasanya ditemukan dengan kombinasi bersama subtipe NA N2) telah menyebabkan wabah dan telah memiliki keturunan yang stabil pada ayam dan unggas darat lainnya, seperti burung puyuh, burung pegar, ayam hutan, dan bebrapa spesies domestik lainnya [82, 85, 157]. Analisis filogenetik dari virus H9N2 dari Asia memberikan kesimpulan bahwa mereka telah bertransmisi dari burung akuatik ke unggas pada beberapa kesempatan [214]. Menariknya, reservoir avian alami dari virus H9 di China belum terindentifikasi, dan virus-virus H9N2 telah terdeteksi hanya pada itik domestik yang kelihatan sehat dari luar dalam kajian surveilans terbatas terhadap LBM dan peternakan di Hong Kong antara tahun 1975 dan 1985 [175, 236]. Pada tahun 1998, tiga virus H9N2 diisolasi dari burung puyuh yang mati pada salah satu peternakan di Hong Kong (40.000 ekor unggas), dimana terjadi wabah penyakit saluran pernafasan yang berlangsung selama sekitar 3 bulan [214]. Isolate-isolat ini adalah bukti pertama adanya virus H9 pada unggas darat di Asia. Bukti-bukti yang tersedia menunjukkan bahwa virus-virus H9 tidak muncul pada ayam di China sampai pada awal tahun 1990-an [82]. Wabah pertama LPAI H9N2 di China Daratan tercatat di Provinsi Guangdong, yang berlangsung dari November 1992 sampai Mei 1994, melanda 17 peternakan ayam dan dua pternakan unggas langka. Angka kematian pada ayam broiler adalah 10-40%, dan tingkat bertelur menurun sampai 14-75% [258]. Sampai akhir tahun 1990-an. Virus H9N2 telah terdeteksi di sebagian besar provinsi di China, terkait dengan aktivitas LBM di sepanjang tahun, dan dengan angka insidensi yang bervariasi dari 0.2% sampai hampir 5% tergantung pada pasar dan waktu tertentu disepanjang tahun [41, 141, 152, 164, 304, 305]. Sejak saat itu, virus LPAI H9N2 telah memperluas cakupan geografisnya. Kombinasi perdagangan unggas secara legal maupun illegal, juga kemungkinan spillover dari burung liar, telah berkontribusi pada menyebarnya virus H9N2 melewati batas negara. Virus LPAI H9N2 keturunan Eurasia ini telah dihubungkan dengan wabah-wabah yang terjadi dai tetap menjadi endemic di pasar-pasar unggas hidup (LBM) dan wilayah operasional perunggasan komersial di banyak negara, meliputi diantaranya Bangladesh, China, Mesir, Jerman, Israel, India, Iran, Lebanon, Pakistan dan Vietnam [13, 15, 48, 51, 92, 114, 187, 189, 190, 196, 207, 232, 259, 271]. Harus dicatat bahwa wabah H9N2 yang terjadi di Jerman tahun 2013-2014 dan di Polandia tahun 2013 bukanlah disebabkan oleh virus yang berasal dari populasi unggas di Asia, berdasarkan pada temuan kurangnya hubungan sekuens gen antara HA dan NA dari isolate Polandia dan virus-virus yang bersirkulasi di wilayah Timur Tengah ataupun Asia Timur Jauh [243, 244]. Analisis filogenetik menunjukkan bahwa virus LPAI H9N2 yang berasal dari Kalkun mewakili masuknya virus dari reservoir burung liar kedalam populasi unggas secara independen, beredar dari tahun 2006 sampai 2013.
Gambar 11.1 Pohon filogeni HA H9 menunjukkan grup-grup filogenetik utama. Analisis filogenetik dilakukan menggunakan alat analisis online yang tersedia pada Influenza Research Database (www.fludb.org). Pohon yang tidak berakar ini dibuat menggunakan perangkat lunak Archaeopteryx seperti yang dideskripsikan dalam referensi [337]. Penandaan dan pemberian warna-warna ditambahkan dengan menggunakan perangkat lunak Powerpoint (Microsoft.Inc.). Lihat bagian Plate untuk melihat representasi warna-warna yang disajikan pada gambar ini.
Faktor yang mungkin paling penting dalam berkontribusi terhadap efektivitas penyebaran virus-virus H9N2 berhubungan dengan banyaknya mode transmisi virus pada unggas, meliputi droplet, fomite, pakan dan air. Di dalam setting laboratorium, dan juga seringkali juga di lapangan, virus-virus H9N2 tidak menyebabkan gejala klinis yang jelas pada unggas, meskipun virus ini dapat bereplikasi dalam level yang tinggi di saluran pernafasan. Beragam penelitian telah menunjukkan bahwa spesies unggas yang berbeda memiliki kerentanan yang bervariasi terhadap virus LPAI. Burung puyuh Jepang tampaknya lebih rentan dibanding ayam dan kalkun, tetapi hanya pada kalkun gejala klinis perrnafasannya dapat teramati [6, 12, 59, 99-102, 128, 169, 192, 198, 214, 237, 239, 274, 296].
Saat wabah H9N2 terjadi pada unggas, biasanya dihubungkan dengan infeksi sekunder bakterial yang kadang berakibat pada angka kematian dan angka kesakitan yang tinggi [23, 109, 114, 118, 126, 197, 199, 237, 144]. Pada saat dilakukan nekropsi, burung-burung yang terinfeksi umumnya menunjukkan lesi konsolidasi pulumonari lokal dari lubang-lubang udara yang tertutup eksudat mengkeju, petekie pada tenggorokan, trakea dan/atau saluran usus, dan sumbatan mukus pada bronkus dan trakea [198, 237]. Menariknya, virus-virus H9N2 kadang-kadang juga ditemukan ber-ko-infeksi dengan virus HPAI H5N1 pada unggas yang tampak sehat dari luar di flok-flok unggas di Mesir [9, 185]. Hasil observasi semacam ini menandai dampak virus H9N2 disebabkan oleh kemampuannya dalam menyamarkan dan mempromosikan transmisi virus HPAI pada unggas. Vaksinasi yang dilakukan untuk mencegah penyakit, terutama dalam bentuk virus inaktif, efektif terhadap virus H9N2 akan tetapi telah gagal dalam mencegah penyebarannya. Setidaknya ada satu virus H9N2 dari keturunan Amerika Utara yang berhubungan dengan vaksin dideteksi juga di China (Gambar 11.1), meskipun tidak diketahui aoakah virus tersebut akhirnya telah punah semenjak itu. Karena virus LPAI H9N2 tidak dimasukkan kedalam daftar penyakit lintas negara yang harus dilaporkan oleh OIE, dan sistem surveilans yang dipergunakan untuk mendeteksi keberadaan virus-virus ini juga sering kali tidak tersedia, cakupan geografisnya dan akar endemisnya mungkin lebih signifikan dari yang telah diketahui sebelumnya.
Virus-virus H9N2 dari keturunan Eurasia telah dibagi ke dalam tiga sub-garis turunan, yaitu GI-like, Y280-like (yang dikenal juga sebagai Beijing/94-like), dan Korean-like (Gambar 11.1) [85]. Virus-virus dalam sub keturunan G1-like telah ditemukan di wilayah Asia dan Eropa, sementara Y280-like dan Korean like tampaknya lebih cenderung tetap berada di Asia [51, 69]. Studi filogeni yang lebih baru menunjukkan pembagian sub-garis yang lebih jauh lagi diantara keturunan G1-like dan Y280-like [69, 156]. Analisis filogeografikal dan evolusi dependensi hospes secara ekstensif telah menyarankan nama-nama clade alternatif yang lebih kompleks untuk virus-virus H9N2 [42, 47, 69, 233, 259], akan tetapi topologi umum dari pohon filogeninya secara umum telah ditunjukkan seperti dalam Gambar 11.1.
Pada saat ini, spillover virus LPAI H9 yang telah beradaptasi terhadap unggas domestik kedalah populasi burung liar telah terdokumentasi di China, akan tetapi belum jelas apakah burung-burung tersebut ataupun spesies avian atau mamalia lainnya telah menjadi hospes permanen [321, 333]. Burung Plateau Pika dari Danau Qinghai di China telah ditemukan memiliki serum antibodi terhadap H9N2m dan juga rentan terhadap infeksi eksperimental dengan virus H9N2 prototipikal, akan tetapi tidaklah jelas apakah mereka mewakili vektor bonafide dalam persebaran virus-virus ini dalam keadaan alaminya, atau apakah mereka juga hanya terpapar, terinfeksi dan menjadi hospes akhir. Hanya ada sedikit subtipe virus H9 yang telah ditemukan pada burung liar Eurasia yang tidak termasuk kedalam keturunan yang juga ditemukan pada unggas domestik, dan bukti kelangkaan terjadinya reassortment antar benua diantara virus-virus H9N2 yang berasal dari keturunan Amerika Utara dan Eurasia telah ditemukan di China [336].
Resiko yang dimiliki virus-virus influenza A H9N2 terhadap Kesehatan Masyarakat
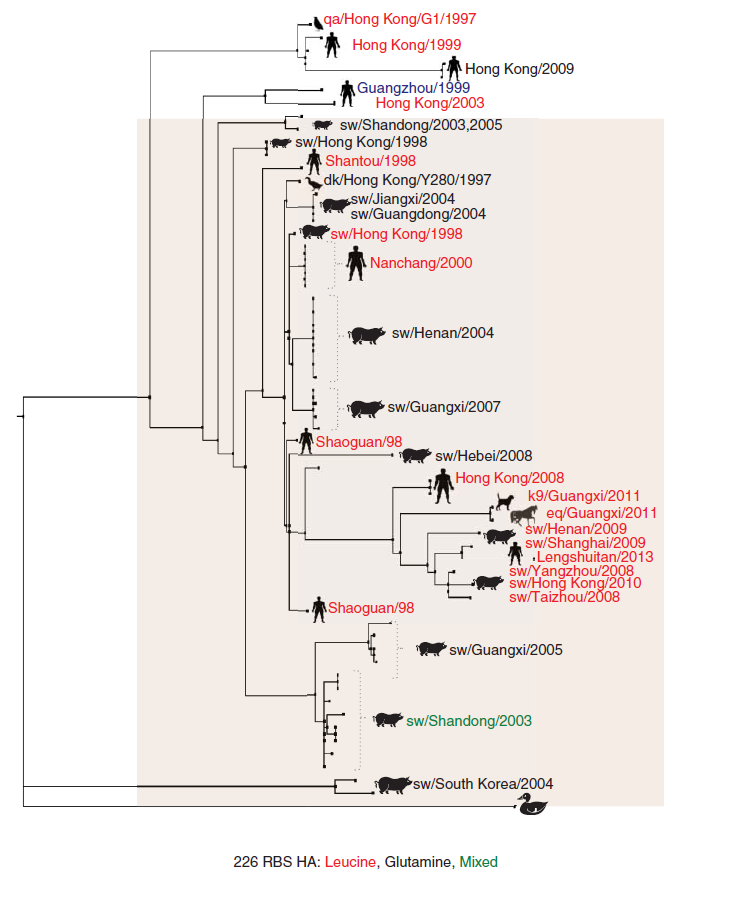
Infeksi virus LPAI h9n2 pada manusia dan babi telah dilaporkan, dengan semua kasus yang terjadi terbatas di Asia (Gambar 11.2). Dua isolate awal manusia virus H9N2 diambil pada tahun 1999 di Hong Kong dari dua pasien yang melaporkan gejala pernafasan ringan [209]. Studi menunjukkan bahwa isolate-isolat ini secara genetic dan antigenik memiliki hubungan dengan keturunan G1-like [147]. Isolate-isolat selanjutnya yang didapat dari kasus sporadic pada manusia selama ini berasal dari keturunan G1-like dan Y280-like [26, 27, 40, 79, 84, 226]. Seluruh kasus pada manusia yang telah dilaporkan sampai saat ini (lebih dari selusin) bersifat ringan dan sembuh sendiri tanpa komplikasi klinis atau infeksi lanjutan, kecuali pada satu kasus positif H9N2 pada pasien kelainan imunitas dengan sejarah penyakit bone-marrow-transplant chronic-graft-versus-host dan penyakit bronchiolitis obliterans yang melanjut kepada kegagalan pernafasan [40]. Jumlah kasus pada manusia yang relative rendah tampaknya karena virus didapatkan melalui kontak langsung dengan burung yang terinfeksi, dan sampai saat ini tidak ada bukti adanya infeksi antar manusia [276]. Pada tahun 1998, virus H9N2 dari babi pertama kali diisolasi di Hong Kong. Infeksi pada babi, tanpa adanya gejala klinis yang jelas, tampaknya jauh lebih umum daripada dari infeksi pada manusia [45, 46, 225, 301]. Infeksi pada babi juga umumnya disebabkan oleh virus Y280-like. Terdapat juga bukti bahwaa virus-virus H9N2 dari keturunan Korea-like telah menginfeksi babi [45, 225, 307]. Dibawah kondisi percobaan, beberapa isolate juga telah dilaporkan menunjukkan patogenitas yang meningkat pada mencit tanpa menimbulkan adaptasi [21, 52, 107, 143, 148, 158, 159, 225, 268, 285, 298, 299, 328]. Pada tahun 2011, infeksi virus H9N2 pada anjing (dan salah satu isolat dari kuda) terdeteksi di Guangxi, China [7, 257, 324]. Pada sebuah studi longitudinal, sampel serum anjing dari lokasi yang sama menunjukkan tingkat seropositif yang tinggi secara signifikan terhadap virus H9N2 yaitu 20.21% di tahun 2010, 28.98% di tahun 2011, dan 44,85% di tahun 2012 [257]. Harus dicatat bahwa studi serologis yang semacam ini haruslah disertai dengan perbandingan menggunakan sampel serum dari wilayah-wilayah dimana virus H9N2 ataupun virus-virus lainnya yang telah beradaptasi pada unggas tidak atau belum ditemukan, dalam rangka mengiliminasi hasil-hasil positif palsu. Secara eksperimental, virus H9N2 dari anjing mampu untuk menginfeksi anjing dan kucing, namun transmisinya hanya terbatas pada kucing [324]. Karena itulah mungkin bagi anjing dan mamalia lainnya yang umumnya dapat ditemukan di pasar-pasar bertindak sebagai hospes perantara yang mampu memperluas sebaran hospes virus H9N2 [7]. Pada beberapa tahun terakhir, virus LPAI H3N2 di Korea dan China dan virus AI H3N8 pada kuda di Amerika Serikat telah beradaptasi pada anjing, menambahkan lagi hospes potensial baru yang dapat bertindak sebagai wadah percampuran virus yang dapat memunculkan varian AI baru [95, 130, 142]. Faktanya, virus reasortant anjing/manusia telah diisolasi dari anjing di Korea selama surveilans rutin [247], menunjukkan bahwa anjing memang dapat bertindak sebagai hospes perantara untuk virus influenza.
Studi serologis pada manusia menunjukkan bahwa ditemukan level paparan yang tinggi pada manusia yang merupakan pekerja peternakan unggas komersial dan pekerja pasar unggas hidup (LBM) dan pada individu yang berkontak dengan unggas yang terinfeksi H9N2. Penelitian di Kamboja, Mesir, India, dan Vietnam telah mengungkap signifikansi paparan virus H9N2 kepada manusia [22, 108, 160, 208, 277, 332]. Jumlah sampel seropositive pada manusia yang diekspresikan sebagai presentasi virus H9N2 adalah sekitar 1-10% (dan bahkan sampai setinggi 40% pada salah satu studi di Iran), terkadang dengan batasan yang cukup jelas antara populasi dalam resiko (yang berkontak dengan unggas) dan populasi pada umumnya. Akan tetapi, diperlukan perhatian dalam hal ini, karena beberapa studi lainnya telah melaporkan bahwa antibody yang bereaksi silang terhadap H2 mungkin dapat menjelaskan meningkatnya jumlah antibody yang bereaksi terhadap H9N2, terutama untuk orang-orang yang lahir sebelum tahun 1968 [195, 254]. Secara keseluruhan, studi-studi serologis haruslah memperhatikan bahwa reaksi silang sebanyak beberapa derajat akan ditemukan pada beberapa individu yang memiliki sejarah infeksi influenza sebelumnya dan atau yang telah divaksinasi sebelumnya.
Gambar 11.2 Pohon filogeni HA H9 menunjukkan hubungan filogenetik yang besar dengan virus-virus H9N2 yang diisolasi dari berbagai spesies hewan. Analisis dan penyesuaian filogeni telah dilakukan seperti yang telah dideskripsikan pada keterangan di atas. Posisi 226 pada situs pengikatan reseptor HA H9 dengan leusin ditunjukkan dengan warna merah, dengan glutamin ditunjukkan dengan warna hitam, dan dengan beragam populasi virus yang membawa leusin dan atau glutamin ditunjukkan dengan warna hijau. Perhatikan bahwa isolate virus tunggal dari kasus pada manusia dengan methonin 226 ditunjukkan dengan warna biru. Kotak berwarna ungu muda merepresentasikan virus-virus dari keturunan G1, dan kotak berwarna coklat muda merepresentasikan virus-virus dari keturunan Y20. Lihat bagian Plate untuk melihat representasi warna-warna yang disajikan pada gambar ini.
Reasortan H9N2 homosubtipik dan heterosubtipik telah diisolasi dari banyak spesies unggas pada banyak pasar unggas hidup (LBM) di seluruh Asia [3, 45, 284, 314, 316]. Virus-virus H9N2 di Asia telah menjadi donor dan resipien segmen gen yang efisien dari virus AI burung liar dan unggas domestik. Kemampuan virus H9N2 yang luar biasa dalam melakukan reassortment telah mengakibatkan banyaknya subtipe influenza pada unggas, beberapa memiliki cakupan hospes yang sangat luas. Subtipe kombinasi HA dan NA yang baru telah muncul dengan gen-gen internal yang berasal dari virus-virus H9N2. Beberapa dari virus-virus ini telah menunjukkan kemampuan untuk menginfeksi mamalia, termasuk manusia. Daftar subtipe-subtipe virus yang umumnya ditemukan pada unggas, terutama pada pasar unggas hidup (LBM) di China dan Asia Tenggara, meskipun ekstensif juga mungkin tidak lengkap dikarenakan kurangnya data surveilans. Harus ditekankan bahwa sistem surveilans hewan pada negara-negara yang kurang berkembang tidak memiliki sumber daya yang cukup untuk melacak keberadaan virus LPAI, dan karena itulah virus-virus tersebut tetap tidak terlaporkan. Untuk menunjukkan signifikansi virus LPAI H9N2 secara penuh perlu disebutkan peranan mereka sebagai donor segmen gen internal kepada virus HPAI H5N1 yang muncul pada unggas di tahun 1996-1996 dan kemudian muncul kembali di tahun 2003-2004. Virus HPAI H5N1 menyebar ke negara-negara ain di Asia, Afrika, dan sebagian Eropa, dan telah menyebabkan infeksi pada manusia [228]. Pada kasus HPAI H5N1 yang telah terkonfirmasi pada manusia, tingkat letalitasnya sangat luar biasa, yaitu di angka sekitar 60%. Antara 2003 dan Juli 2014, organisasi kesehatan dunia WHO telah melaporkan 667 kasus HPAI H5N1 pada manusia dengan 393 kasus fatal (www.who.int/influenza/human_animal_interface/acian_influenza/en/). Negara-negara seperti Kamboja, China, Mesir, Indonesia dan Vietnam, dimana virus HPAI H5N1 tetap endemic, terus melaporkan infeksi pada manusia meskipun sudah jarang terjadu [170, 293]. Terkini, infeksi virus LPAI pada manusia dari subtipe H7N9 dan H10N8 juga telah dilaporkan [30, 270, 326]. Kedua virus ini berbagi segmen gen internal yang sama yang berasal dari virus H9N2 yang telah beradaptasi pada unggas domestik yang telah bersirkulasi di China [64, 72, 219]. Hanya dua kasus infeksi H10N8 yang telah dilaporkan, keduanya di provinsi Jiangzi, China [34, 327]. Salah satu dari kasus ini adalah wanita berumur 73 tahun yang mengalami demam, masuk rumah sakit pada tanggal 30 November 2013, dan meninggal enam hari berikutnya karena kerusakan multi organ. Wanita tersebut telah mengunjungi pasar unggas hidup (LBM) empat hari sebelum gejala muncul. Virus penyebabnya mengandung campuran segmen gen PB2 yang mengkode E627 dan K627, dan bersifat sensitive terhadap hambatan neuraminidase [34].
Infeksi virus baik LPAI maupun HPAI subtipe H7 pada manusia telah dilaporkan meskipun jarang, dan sebagian besar dari kasus-kasus ini dihubungkan dengan kasus konjungtivitis ringan [2, 17, 29, 56, 66, 163, 275]. Sampai tahun 2013, hanya ada satu pengecualian yang terjadi tahun 2003, saat terjadi wabah HPAI H7N7 pada peternakan unggas komersial di Belanda. Virus HPAI H7N7 juga terdeteksi pada 88 orang yang menunjukkan gejala konjungtivitis atau gejala pernafsan ringan, dan pada satu orang yang meninggal karena pneumonia dan sindrom pernafasan akut [66]. Seluruh kasus pada manusia tersebut telah berkontak dengan unggas selama terjadinya wabah, dan dicurigai adanya 30 transmisi manusia ke manusia, akan tetapi tidak terdapat transmisi menerus ke orang yang selanjutnya [54, 122]. Yang lebih menjadi perhatian adalah situasi terkini terkait virus LPAI subtipe H7N9 di Asia. Antara Maret 2013 dan Oktober 2014 [28, 191]. Sebanyak 453 kasus LPAI H7N9 yang terkonfirmasi di Laboratorium, termasuk 175 kematian yang disebabkannyam telah dilaporkan kepada WHO (www.who.int/influenza/human_animal_interface/influenza_h7n9/en/). Kasus-kasus ini telah dilaporkan dari China bagian timur, dan diasosiasikan dengan aktivitas di pasar unggas hidup (LBM) selama bulan-bulan musim dingin di provinsi Anhui, Fujian, Guangdong, Guangxi, Hebei, Hunan, Jilin, Jiangsu, Jiangxi, Shanding, dan Zhejiang, dan di Kotamadya Beijing [33, 105]. Terdapat juga 10 kasus yang dilaporkan di Hong Kong SAR [37, 269], empat kasus di Taipei, Taiwan [31, 146, 162, 173], satu kasus dari pelancong asal China di Malaysia, dan dua kasus dari Wilayah Otonomi Khusus Xinjiang [330], yang berbatasan langsung dengan Rusia, Mongolia, Kazakhstan, Lirgistan, Tajikistan, Afganistan, Pakistan dan India. Pasien yang terkonfirmasi mengalami infeksi virus H7N9 saat masuk rumah sakit menunjukkan salah satu atau lebih gejala klinis yang meliputi demam tinggi, batuk produktif atau non-produktif, nafas pendek, dyspnea dan hipoksia. Hasil pemeriksaan X-Ray menunjukkan penyakit saluran pernafasan bawah, umumnya melaporkan keberadaan peningkatan opasitas/bidang putih, konsolidasi dan infiltrasi cairan. Kasus infeksi berat H7N9 menunjukkan gejala shock sepsis, kegagalan respirasi, sindrom susah bernafas akut, hipoksemia refraktori, disfungsi ginjal akut, disfungsi multiple organ, rhabdomiosis, dan encepalopati. Infeksi sekunder bakteri dan jamur juga telah dilaporkan, kadang-kadang dihubungkan dengan bakteria yang resisten terhadap banyak obat [36, 71, 106, 17, 150, 165-167, 286-288, 295, 310, 313, 319, 322, 330, 331]. Dengan pengecualian sedikit infeksi virus H7N9 yang menunjukkan gejala penyakit saluran pernafasan atas yang tidak kompleks dan demam ringan, baik pada anak-anak maupun orang dewasa, sebagian besar infeksi bersifat parah. Waktu median dari onset munculnya penyakit sampai dibawa ke rumah sakit rata-rata dalam 4,5 hari, dan pasien-pasien ini memerlukan perawatan intensif. Waktu dari onset penyakit sampai kemaian bervariasi dari 7 sampai 20 hari. Steroid sistemik dosis tinggi tampaknya berhubungan dengan meningkatnya resiko perpanjangan shedding virus dan munculnya reisistansi terhadap antiviral [106]. Munculnya virus H7N9 kembali mengingatkan ketidakpastian dari sifat virus AI. Tidak ada bukti aktivitas virus H7N9 yang teramati pada unggas sebelum kasus pada manusia dilaporkan, meskipun virus-virus ini tampaknya telah menyebar di mana-mana pada pasar unggas hidup (LBM) sejak saat itu [63, 116, 127, 249, 320]. Virus-virus H7N9 tampaknya tidak menunjang transmisi manusia ke manusia. Sebaliknya, perlu dicatat bahwa ayam dan unggas lainnya tampaknya tidak menunjukkan gejala klinis yang jelas jika terinfeksi dengan virus H7N9, akan tetapi dapat menyebarkan virus [206]. Saat ayam, burung puyuh, burung merpati, dan berbagai spesies itik diinfeksi secara eksperimental dengan strain H7N9 prototipikal, langsung terobservasi adanya replikasi virus yang signifikan, dengan level transmisi yang bersifat sedang (intermediate) yang berada diantara level transmisi virus patotipikal manusia dan virus-virus AI lainnya [18, 222, 334]. Tidak perduli jenis patotipikalnyauntuk unggas, karakteristik utama infeksi virus H7 pada musang adalah potensi penyebarannya ke otak [19, 115, 263]. Tidak seperti virus AI lainnya, secara umum virus H7 tidak memerlukan saklar pada spesifisitas reseptor dalam rangka menginfeksi manusia, dan mereka dapat menyebabkan infeksi dan dapat ditransmisikan, meskipun hanya parsial, pada model mamalia dengan masih mempertahankan preferensi tempat perlekatan asam sialic a2,3α-Gal yang mirip dengan pada unggas [18, 20, 222, 248, 263, 334]. Karena sumber virus H7N9 masih belum diketahui benar dan belum dapat dikontrol, kasus-kasus pada manusia yang lain mungkin akan muncul [206]. Meskipun jumlah kasus telah menurun secara dramatis pada bulan-bulan musim panas, kasus yang muncul terakhir dilaporkan dari Wilayah Otonomi Khusus Xinjiang Uyghur, yang sebelumnya tidak pernah melaporkan kasus, dan tidak berdekatan dengan wilayah yang sebelumnya terdampak, yang mengindikasikan bahwa virus masih terus bersirkulasi dan mengembangan wilayah geografisnya. Penutupan pasar unggas hidup (LBM), disenfeksi, waktu istirahat, dan pemusnahan unggas telah terbukti efektif dalam mengontrol infeksi pada manusia, akan tetapi tampaknya hanya memberi sedikit dampak dalam membatasi kemunculan Kembali dan penyebaran virus HPAI dan LPAI di Asia [67, 68, 97, 186, 278, 297, 315]. Dinamika yang kompleks dari praktek pertanian terintegrasi dan hubungannya yang sangat dekat dengan system pemasaran hewan hidup menyebabkan pencegahan, penanggulangan dan eradikasi virus LPAI menjadi sebuah tugas yang sangat sulit.
Penting untuk mengenali bahwa penanda molekuler untuk pergantian hospes masih belum terlalu baik untuk virus IA yang berasal dari reservoir alaminya (misalnya dari burung-burung akuatik) kepada unggas. Dengan beberapa pengecualian, sebagian besar upaya diarahkan untuk dapat memahami transmisi virus AI antar spesies dari burung ke mamalia, akan tetapi tidak untuk memahami transmisi antar spesies burung yang berbeda. Juga penting untuk ditekankan bahwa sejumlah besar studi telah dapat mengindetifikasi mutasi yang meningkatkan virulensi virus LPAI (dan HPAI) pada hewan model (tikus), akan tetapi peranan mutasi ini terhadap infeksi pada mamalia lain masih belum jelas.
Hemaglutinin
Situs perlekatan reseptor
Sejumlah besar isolate H9N2 dari unggas mengandung leusin pada asam amino 226 (L226) pada situs perlekatan reseptor (RBS) dari HA (H3, 216 pada HA H9 yang mature), dan menunjukkan affinitas yang lebih besar untuk berikatan dengan reseptor analog dengan asam sialic (SA) yang dihubungkan dengan galaktosa melalui hubungan α2,6 (SA α2,6Gal) [177, 281]. Ini adalah dua karakteristik umum virus AI manusia. Sebaliknya, virus-virus H9N2 yang mengandung glutamin pada asam amino 226 (Q226) pada HA menunjukkan peningkatan preferensi untuk SA α2,3Gal [281]. Tergantung pada kontek tempat perletakan asam aminonya di dalam ataupun berdekatan dengan RBS, virus-virus yang mengandung Q226 juga dapat menunjukkan sensitivitas reseptor berganda [281]. Pengambilalihan L226 pada virus-virus H9N2 telah terjadi pada unggas darat [41, 141, 304], konsisten dengan hasil observasi bahwa baik reseptor SA α2,3Gal dan SA α2,6Gal terdapat pada saluran pernafasan pada beberapa dari spesies ini [70, 83, 119, 282]. Burung puyuh, burung pegar, ayam Mutiara, dan kalkun telah diketahui memiliki kedua tipe reseptor ini pada saluran pernafasan dan pencernaanya, sedangkan itik dan angsa memiliki hampir ekslusif satu tipe saja yakni α2,3Gal [119, 318]. Bukti-bukti yang ada menunjukkan bahwa L226 pada HA virus H9N2 sesekali mendorong infeksi pada mamalia, terutama manusia (Gambar 11.2). Virus-virus yang mengandung L226 tumbuh lebih efisien pada kultur sel epithelial saluran pernafasan manusia (HAE) jika dibandingkan dengan virus-virus yang mengandung Q226 jika dipertahankan dalam media air-liquid interface (ALI) [281]. Akan tetapi, virus-virus yang mengandung Q226 juga telah diidentifikasi dari babi dan sesekali pada manusia (Gambar 11.2). Pada inokulasi eksperimental pada kera Rhesus macaques, infeksi virus H9N2 menimbulkan respons demam dua fase bersama terdeteksinya replikasi virus di saluran pernafasan atas dan bawah [323]. Dalam kondisi eksperimental keberadaan L226 pada HA memungkinkan replikasi dan transmisi yang efektif pada musang dan marmot melalui kontak langsung, meskipun transmisi melalui udara sampai kontak tidak langsung tidak ada sama sekali atau kurang efisien dibandingkan dengan virus AI pada manusia [144, 283]. Musang dianggap sebagai standar emas untuk mempelajari transmisi virus AI manusia, karena hewan tersebut rentan, dan mampu mentransmisikan virus melalui udara, dan menunjukkan gejala klinis yang mirip dengan gejala yang terobservasi pada manusia. Virus-virus H9N2 yang mengandung L266 cocok untuk reassortment dengan virus H3N2 manusia yang bersifat musiman (sH3N2) dan juga dengan virus-virus H1N1 (pdm) penyebab pandemic tahun 2009 [120, 121, 220, 251, 260, 283]. Banyak hasil reassortansi ini menunjukkan peningkatan infektivitas dan meningkatkan kemampuan transmisi melalui kontak langsung pada musang. Pada salah satu studi, salah satu virus reassortan yang memiliki gen permukaan dari virus avian H9N2 dan gen internal dari virus influenza musiman H3N2 manusia (sH3N2) hanya ditransmisikan melalui kontak langsung pada musang. Transmisi melalui udara lebih sedikit meskipun virus reasortan H9N2 avian/manusia menyebabkan gejala sakit klinis dan temuan patologis pada musang dengan cara yang sama dengan virus sH3N2 [238]. Adaptasi lebih lanjut dari pasase berantai pada musang dengan virus reasortan H9N2 avian/manusia menunjukkan hasil perubahan asam amino yang minimal yang sesuai untuk transmisi lewat udara pada hewan-hewan ini. dua perubahan asam amino pada HA H9, yaitu T189A pada wilayah HA1 dan G192R pada wilayah H2, penting untuk transmisi lewat udara [251]. Transfer segmen gen HA ini yang terjadi di “balik layar” pada strain pdm 2009 (H9N1, reasortan 1+7) juga berakibat transmisi lewat udara yang efisien [121]. Kontribusi perubahan-perubahan kecil ini pada HA juga terobservasi pada virus reasortan alternative H9N1 avian/pdm (3+5 dan 5+3) [120], menandai adanya dampak perubahan asam amino yang kecil pada penotipe transmisi virus. Mutasi T189A (179 pada sekuensi HA H9) tidak bersifat untuk hanya pada virus H9 yang telah beradaptasi pada musang. Meskipun sebagian besar isolate alami mengandung threonine, strain virus yang mengandung alanin, asparagin, isoleusin, proline dan serine juga telah terindentifikasi. Isolate H9 alami dengan mutasi pada G192R (182 pada sekuens HA H9) jarang terjadi, dengan hanya satu sekuens telah dilaporkan, sedangkan hanya beberapa saja yang menunjukkan adanya susbtitusi G192D.
Virus HA H7N9 yang menyebabkan infeksi pada manusia di Asia membawa perubahan molekuler yang bersifat prediktif untuk dikenali oleh reseptor SA α2,6Gal yang mirip pada manusia. Mutasi Q226L dan G186V (Penomoran H3 adalah 217 dan 177, pada HA H7 dewasa/mature) menyediakan beberapa level pengenalan pada tempat perlekatan reseptor mirip-manusia. Akan tetapi virus H7N9 tidak dapat bereplikasi dengan baik dalam sel epitel trakea manusia, dan mampu mempertahankan perlekatannya secara signifikan kepada reseptor mirip-avian [53, 235, 267, 300, 306].
Situs pembelahan proteolitik
Jika dibandingkan dengan penanda molekuler, alih-alih alanine, keberadaan serine pada posisi P5 pada situs pembelahan HA H9 (PSRSSR/GL) telah diperlihatkan mampu meningkatkan efisensi pembelahan dan meningkatkan replikasi pada ayam dan mencit [261]. Secara in vitro, virus-virus HA H9 dapat dibelah oleh protease ekstraselular dari saluran pernafasan, seperti protease transmembrane manusia, serine S1 member 2 (TMPRSS2) dan protease mirip tripsin di saluran nafas manusia (HAT) [16]. Serupa dengan itu, HA dari virus H7N9 dikenali dengan TMPRSS2, aktivitas yang tampaknya penting untuk siklus hidup dan pathogenesis virus [227, 265]. Menariknya, HA H9 dengan tempat pembelahan sekuens RSSR/GL atau RSRR/GL dapat dibelah oleh matriptase, sebuah protease yang umum diekspresikan pada banyak epithelia. Matriptase diekspresikan dalam jumlah banyak di ginjal, dan mungkin dapat memberikan sebagian penjelasan nephrotropisme pada beberapa virus H9N2 yang terobservasi pada ayam [16]. Dalam situasi special dimana terdapat hilangnya tempat glikosilasi, HA H9 dapat dibelah oleh furin. Pembelahan oleh furin bersifat atipikal untuk HA dari virus LPAI akan tetapi dapat mempengaruhi kemampuan virus H9N2 untuk menyebar pada unggas [273]. Pertanyaan apakah HA dari virus-virus H7N9 adalah substrat dari protease atau mekanisme yang serupa membutuhkan perhatian lebih.
Neuraminidase
Banyak strains H9N2 membawa delesi pada wilayah cabang NA, yang bersifat tipikal pada virus AI yang telah beradaptasi dengan tropisme respiratori pada unggas [3, 4, 14, 132, 147, 153, 155, 246, 307, 325]. Studi laboratorium telah menunjukkan bahwa pasase berantai isolat virus AI dari burung liar pada ayam ataupun burung puyuh menyebabkan berkembangnya strain yang lebih beradaptasi baik terhadap unggas yang membawa penghapusan tangkai NA [74, 104, 250]. Banyak isolat H9N2 Eurasia yang muncul secara alami dari unggas membawa delesi 3 asam amino dari tangkai NA melibatkan posisi 62-64, yang juga tampaknya meningkatkan virulensinya pada mencit [85, 148, 158]. Delesi 2 asam amino alternatif melibatkan posisi 38-39 terobservasi pad dua dari strain pada manusia di Hong Kong pada tahun 1999 [147]. N2 NA dengan delesi asam amino 38-39 belum menjadi tetap pada populasi virus dan belum dilaporkan lagi sejak tahun 1999, kecuali pada dua virus H9N2 dari Jepang yang diisolasi dari burung Parkit yang diimpor dari Hong Kong [176]. Tidak ada hubungan langsung yang telah terbentuk antara delesi cabang dan transmisi dari unggas ke manusia atau babi. Tidak ada bukti kenaikan resistensi terhadap inhibitor NA (NAI) telah ditemukan pada virus-virus H9N2, dan tidak ada satupun dari 850 sekuens N2 NA yang dianalisis memiliki resistensi substitusi R292K atau H274Y NAI. NA dari virus-virus H7N9 dari kasus pada manusia dan yang berasal dari strain asal yang berawal dari unggas membawa delesi yang melibatkan asam amino 68-72. Setidaknya tiga dari lebih dari 180 sekuens menunjukkan mutasi R292K yang akan membawa resistansi terhadap inhibitor neuraminidase [86]. Mutasi R292K tampaknya tidak mempengaruhi kebugaran dan virulensi virus [86].
Kompleks Polimerase
PB2
Dalam segmen gen internal, protein PB2 yang memiliki mutasi-mutasi mengindikasikan perbaikan replikasi pada mamalia (E627K atau D701N) [93, 94, 129, 168, 256] bersifat tidak umum diantara virus-virus H9. Sebagian besanya memiliki profil tipikal avian-like E627, D701. Hanya empat (dari lebih dari 700) strains H9N2 yang muncul secara alami yang telah dideskripsikan sampai saat ini menunjukkan mutasi K627 pada PB2, dan tidak ada bukti bahwa mutasi itu telah bersifat tetap pada populasi virus. Beberapa studi adaptasi pada burung puyuh dan mencit juga menunjukkan bahwa virus-virus H9 dengan segmen PB2 mengandung K627, yang mengindikasikan peningkatan replikasi virus pada saluran pernafasan bawah [21, 104, 143, 148, 158, 159, 225, 285, 298, 299, 328]. Menariknya, tidak ada dari isolate H9N2 yang muncul secara alami yang didapat dari babi atau manusia yang mengandung mutasi PB2 E627K. Sebuah kluster virus H9N2 dari Israel, yang kemudian menyebar ke Mesir, mengandung mutasi PB2 E627 [1, 8, 9, 13, 48, 49, 216, 217], yang juga muncul pada dua isolat yang didapat dari ayam di Hong Kong pada tahun 2011. Mutasi PB2 E627 tampaknya sudah bersifat tetap pada virus-virus H9N2 yang bersirkulasi di Israel dan Mesir, akan tetapi tidak di Hong Kong. Mutasi tidak biasa pada posisi 627 di PB2 telah dilaporkan di dua isolat ayam yang berbeda, satu mengandung susbtitusi E627G [152] dan satu lagi dengan mutasi E627A [49]. Sebalinya, sepertiga (63 dari 186) dari sekuens PB2 H7N9 yang tersedia mengandung substitusi E627K. Isolat H7N9 dengan mutasi E627K biasanya didapat dari manusia akan tetapi tidak dari unggas [183]. Mutasi PB2 D710 terdapat pada satu isolat H9N2 yang diambil dari pasien yang mengalami kelainan sistem imun, yang akan menunjukkan bahwa dibawah kondisi tertentu virus H9N2 mungkin akan mendapatkan mutasi yang khusus untuk beradaptasi pada manusia [40]. Di dalam laboratorium, mutasi tambahan PB2 pada strain H9N2, yaitu D253N dan Q591K telah dihubungkan dengan meningkatnya akivitas polymerase, meningkatnya replikasi pada sel ALI HAE, meningkatnya ekspresi TNF-α pada makrofag manusia dan meningkatnya patogenitas pada mencit [184]. Mutasi PB2 D253N dan Q591K sangat tidak biasa diantara virus LPAI H9, dan mungkin merefleksikan seleksi virus menunjukkan fenotipe plat besar pada sel kultur jaringan (MDCK). Analisis lebih lanjut terhadap sekuens PB2 dari H9 dan H7N9 telah menunjukkan tidak ada isolat natural mutasi D253N, sedangkan mutasi Q591K hanya ditemukan pada tiga isolat virus H9N2 avian dan enam virus H7N9. Menariknya, mutasi PB2 D253N ditemukan pada virus reasortan H9N1 avian/manusia setelah adaptasi berantai dan transmisi udara pada musang, menunjukkan adanya potensi adanya peranan dalam adaptasi virus kepada mamalia [120]. Pada posisi 591, mutasi Q591R juga ditunjukkan dapat meningkatkan virulensi virus prototipikal HPAI H5N1 pada mencit [308]. Mutasi Q591R juga memodulasi virulensi virus pdm H1N1 2009 [182, 308]. Pada PB2, posisi 591 pada virus H9 dan H7 umum ditemukan baik glutamin maupun leusin, akan tetapi tidak ada arginin yang terdeteksi. Pada sebuah studi yang berbeda, mutasi M147L di PB2, kombinasi E627K, telah menunjukkan peningkatan patogenitas pada mencit [285]. Posisi PB2 147 pada strain H9 menunjukkan beberapa plastisitas pada strain yang membawa metionin, isoleusin, treonin, atau valin. Akan tetapi, leusin belum terdeskripsikan pada strain yang terjadi secara alami. PB2 dari strain H7N9 mengandung isoleusin pada posisi 147. Baru-baru ini, mutasi PB2 F404L telah menunjukkan peningkatan aktivitas polymerase dan virulensi pada mencit pada virus H9N2, juga strain HPAI H5N1 dan pdm 2009 [158, 159]. Segmen PB2 yang ada baik pada strains H9N2 dan H7N9 menunjukkan konservasi penilalanin dalam level tinggi pada posisi 404. Mutasi alternatif yang didapat selama adaptasi reasortan virus H9N1 dan H9N2 avian/manusia pada musang memproduksi mutasi pada posisi T581 dan L3741 [121, 251]. PB2 T58, berlokasi pada area perlekatan PB1 dan NP yang bertumpang tindih satu sama lain, terkonservasi dengan sangat tinggi diantara virus H9, hanya beberapa isolat saja yang memiliki alanin (A58), akan tetapi tidak ada yang memiliki isoleusin. Pada studi yang berbeda dimana virus HPAI H7N7 diadaptasikan pada musang, mutasi serupa pada treonin dan isoleusin terobservasi pada area perikatan PB1/NP yang sama dari PB2, akan tetapi pada posisi berbeda, yaitu posisi 81 (T811) [263]. Mutasi PB2 T811 umum ditemukan pada virus H9, yang juga mengakomodasi metionin dan alanin. Begitu juga mutasi PB2 L3741 bersifat umum diantara virus-virus H9. Terakhir, mutasi PB2 A707T telah terobservasi pada virus H9N1 alternatif unggas/manusia yang ditransmisikan melalui udara pada musang [120], akan tetapi perubahan yang semacam ini tidak biasa untuk virus H9, dengan sekuens yang memiliki alanin atau serine, akan tetapi tidak memiliki threonine. Mutasi-mutasi lain pada PB2 telah menunjukkan kemampuan memodulasi virulensi virus H1N1 pdm 2009 pada mencit meliputi E158G/A, H357N, I504V, T5881, dan G90S [110, 111, 161, 224, 329, 335]. Penting untuk diperhatikan bahwa virulensi virus pdm H1N1 2009 sangat dipengaruhi oleh adanya perubahan pada segmen gen permukaan, dan karena itulah dampak mutasi-mutasi lainnya tampaknya lebih spesifik kepada strainnya, dan merupakan hasil dari adaptasi virus secara langsung pada mencit [311]. Dalam PB2 pada virus H9N2 dan H7N9, varian E158, H357, dan V504 adalah varian yang sangat terlindungi dari kerusakan, sementara posisi 588 berkorespondensi terutama sekali dengan alanin. Mutasi G590S tidak terlihat pada virus H9, akan tetapi 8% dari virus H7N9 memang memperlihatkan substitusi itu. Signifikansi perubahan epistatik untuk transmisi antar spesies dan adaptasi virus LPAI pada mamalia masih belum terjelaskan.
PB1
Tidak ada hubungan langsung yang ditemukan untuk mutasi PB1 yang terlibat dalam cakupan hospes virus H9N2 dan H7N9. Sampai saat ini dari keseluruhan sekuens yang dianalisis sampai saat ini tidak ada yang mengandung mutasi H99Y, akan tetapi banyak dari mereka yang mengandung mutasi 1368Y, yang jika keduanya digabungkan telah terbukti sebagai faktor penting yang mempengaruhi transmisi udara strain virus HPAI H5N1 pada musang [98, 149]. Mutasi L473 dan L598 yang terdapat pada virus HPAI H5N1 dapat meningkatkan virulensi strain A/WSN/1933 yang telah beradaptasi pada laboratorium pada mencit. Strain ini membawa mutasi PB2 E627 [302]. PB1 V473 mendominasi di antara strain H9 dan H7N9, akan tetapi P598 tidak biasanya ditemukan, dan hanya tiga strain H9N2 yang muncul secara alami yang mengandung mutasi L598P (dari lebih dari 600 sekuens). Strains H9N2 dari Asia, seperti juga virus LAPI dan HPAI dari wilayah yang sama, menunjukkan beberapa derajat variasi pada ujung terminal N dan C dari protein PB1. Ujung terminal dari PB1 mengandung domain perlekatan PA, yang paling terjaga baik diantara virus avian influenza [96, 212, 213]. Efek mutasi pada wilayah ini masih belum diketahui. Sejumlah besar virus LPAI Asia, termasuk strain-strain H9N2 dan H7N7, mengkode asam amino tambahan pada ujung terminal C, yang mengandung tempat perlekatan PB2 [78, 202, 218, 221] dan memiliki sekuens 753-LRRQKQ-758 atau 753-LGRQGK-758 daripada sekuens tipikal 753-LRRQK-757. Variasi minor dari tiga konsensus sekuens ini juga ditemukan. Mutasi alternatif pada PB1, D120N dan D439E dan S161N telah terobservasi pada virus H9 yang telah beradaptasi untuk menyebar melalui udara pada musang [120, 121, 251]. Signifikansi biologis dari adanya polimorfisme seperti itu masih belum tereksplorasi.
PA
Pada PA, dua mutasi telah secara independent memiliki perasan dalam menimbulkan virulensi pada hewan model tikus. Subtitusi PA K185R telah ditunjukkan menurunkan virulensi virus HPAI H5N1 [61], sementara mutasi T91 telah ditunjukkan meningkatkan virulensi virus LPAI H6N1 [38]. Sebagian besar strain H9 dan seluruh strains H7N9 memiliki segmen gen PA yang mengandung R185 dan T97. Posisi T97 menunjukkan beberapa variasi pada virus H9N2, dengan beberapa strain mengandung antara A97 atau N97, akan tetapi tidak 197. Mutasi tambahan, K26E, dalam situs endonuclease PA telah dideskripsikan pada salah satu virus H9N1 avian/manusia yang telah disebutkan sebelumnya [120]. Akan tetapi analisis sekuens PA dari virus H9 dan H7N9 menunjukkan bahwa E26 lebih dipilih daripada K26, dan karena itulah mutasi K26 mungkin mewakili seleksi dalam mencari residu yang lebih stabil yang dapat meningkatkan keseluruhan aktivitas polymerase yang tidak tergantung dengan kondisi pada lingkungan hospes. Mutasi tambahan pada PA telah ditunjukkan mampu memodulasi virulensi pada mencit dan/atau juga memodulasi aktivitas polymerase, terutama dalam konteks strain pdm H1N1 2009, meliputi F35L, A36T, T851, P224S, L295P, E298K, L336M, dan I550L [25, 111, 161, 224, 231, 262, 311, 335]. Juga harus dicatat bahwa beberapa mutasi ini tampaknya tidak berpengaruh pada infeksi virus AI pada manusia. Mungkin yang lebih relevan adan temuan yang mengungkap bahwa pada virus H7N9 yang telah menginfeksi manusia, mutasi PA S409N tampaknya meningkatkan viral fitness [309]. Meskipun baik serin maupun asparagin 409 terdapat pada virus-virus H9N2 dan H7N9, serine tampaknya lebih dipilih, yang akan menyebabkan lebih sedikit kesempatan untuk menginfeksi mamalia. Signifikansi polimorfisme ini pada PA yang mempengaruhi cakupan hospes dan virulensi memerlukan studi yang lebih jauh lagi.
Gen Nukleoprotein
Meskipun NP telah dikenali dari awal sebagai faktor cakupan hospes, perubahan pada NP yang diasosiasikan dengan transmisi antar spesies masih belum dikenali dengan jelas. NP A184K membawa ke meningkatnya virulensi prototipikal infeksi virus HPAI H5N1 pada ayam, berhubungan dengan meningkatnya titer virus dan meningkatnya level nitrit oksida pada jaringan, demikian juga regulasi IFN-α, IFN-γ, Mx1, dan iNOS, diantara yang lain-lainnya [289]. Serupa, pada berbagai studi yang berbeda terhadap mutasi NP I109T meningkatkan neurotropisme virus HPAI H5N1 pada ayam [264]. Baru-baru ini, subtitusi N52Y pada NP telah ditunjukkan meningkatkan sensitivitas dari virus prototipikal H7N9 kepada Mx pada manusia, mengindikasikan bahwa residu ini adalah penentu resistensi Mx pada manusia [223]. Analisis dari lebih dari 800 sekuens NP dari virus H9N2 dan H7N9 mengungkapkan derajat konservasi isoleusin dan lisin yang tinggi pada posisi 109 dan 184. Hanya varian minor yang ditemukan pada virus H9N2, dengan posisi 109 membawa serin, threonine, atau valin, dan posisi 184 membawa arginin atau methionine. Akan tetapi, NP posisi 52 menunjukkan lebih banyak variasi sekuens, dengan virus-virus yang memiliki asparagin, glutamin, histidine atau tirosin (satu isolate H9N2 dan satu isolate H7N9 mengandung serine). Tidak ada bukti langsung bahwa mutasi NP mempengaruhi transmisi antar spesies yang telah dijelaskan.
Gen Protein Matriks
Strain H9N2 dan H7N9 yang resisten terhadap adamantane adalah umum dikarenakan adanya mutasi S31N pada pompa saluran proton M2, karakteristik yang khas pada fenotipik tersebut. Mutasi V27A yang berhubungan dengan situs kedua pada M2 jarang ditemukan yang terlibat dalam resistansi adamantane.
Mutasi pada M1 yang terkait dengan cakupan hospes atau virulensi virus H9N2 dan H7N9 belum diketahui. Mutasi M1 T139 berhubungan dengan meningkatnya virulensi
Gen Protein non-struktural
Salah satu dari protein virus influenza yang paling banyak dipelajari adalah NS1, yang mengandung N-terminal RNA-binding domain dan C-terminal effector domain [55, 87, 88, 145, 172]. Pada burung-burung liar, dua alel segmen gen NS telah dijelaskan, yakni A dan B. Alel A adalah yang telah dijelaskan secara ekstensif, karea ditemukan pada virus pada manusia dan mamalia lainnya. Protein NS1 bervariasi panjangnya, dengan strain yang mengkode 217 protein asam amino, protein dengan 237 asam amino, sedikit strain mengkode protein dengan panjang intermediate, dan beberapa mengkode 237 protein asam amino. Efek keseluruhan dari NS1 adalah untuk menghadapi respons antiviral yang ditimbulkan oleh hospes setelah infeksi [11, 123, 125, 253, 266]. Detail dari interaksi faktor hospes NS1 dan modulasi aktivitas-aktivitas ini melebihi cakupan Bab ini. Virulensi yang disebabkan NS1 telah dihubungkan dengan keberadaan dari setidaknya salah satu dari marker berikut: P42S, E92D, 1106M, dan sekuens C-terminal ESEV/EPEV. Lebih dari 1000 sekuens NS1 protein telah tersedia untuk virus H9, Sebagian besarnya mengandung S42 dan Sebagian kecilnnya mengkode A42, akan tetapi tidak ada yang mengkode proline. Terdapat 200 sekuens dengan panjang penuh yang tersedia untuk segmen NS dari H7N9, kesemuanya menkode S42. NS1 E9 sangat tidak biasa untuk virus H9N2 dan H7N9, dan Sebagian besar strain mengkode S42. NS1 I106M mengembalikan perlekatan dengan CPSF30 dan memodulasi virulensi pada mencit pada virus H7N9 asal Asia [10], segmen gen internal yang diturunkan dari virus H9N2. Di alam, NS1 dengan I106 atau M106 biasa ditemui, dan karena itulah bisa dispekulasikan bahwa virus H9N2 dan H7N9 yang ditemui secara alami di lingkungan berbeda potensinya dalam menginfeksi dan/atau menyebabkan penyakit pada mamalia. Sekuens ESEV/EPEV berkorespondensi domain perlekatan PDZ [76, 88]. Faktor selular dengan motif PDZ telah diidentifikasi yang berinteraksi dengan NS1 melalui domain ESEV/EPEV, meliputi Scribble, D1g1, PDlim2, dan MAGI-1, yang pada akhirnya berakibat pada modulasi interferon dan signal apoptotik pada sel yang terinfeksi virus influenza [62, 76, 124, 151, 317]. Strains H9N2 dan H7N9, seperti juga banyak virus AI lainnya, mengkode 230 asam amino protein NS1 yang mengandung sekuens ESEV/EPEV. Motif ESEV/EPEV telah ditunjukkan sebagai faktor virulen pada virus H1N1 yang beradaptasi pada tikus [112], akan tetapi memiliki efek terbatas pada virus HPAI H5N1 pada mencit [252]. Kira-kira 50% dari sekuens H9 NS1 yang tersedia merepresentasikan versi truncated dari 217 asam amino, yang konsisten dengan notion bahwa trucations Dampak dari replikasi dan penyebaran virus AI pada burung [252].
Perubahan-perubahan lain
Diantara komponen-komponen protein kecil dari virus AI yaitu NS2/NEP [200] dan PB1-F2 [35], peranan-peranan telah ditugaskan untuk virulensi yang dicapai melalui modulasi berbagai proses viral dan seluler. Produk-produk protein virus lainnya juga telah ditemukan, meskpun fungsinya belum dapat terjelaskan, terutama didalam konteksi terkain virus AI. Protein-protein ini yaitu PB1-N40, PA-X, PA-N155, PA-N182, M42 dan NS3 [65, 113, 188, 234, 279, 294].
Fungsi NS2/NEP yang paling terkarakterisasikan adalah sebagai faktor viral yang bertanggungjawab dalam eksportasi kompleks M1-vRNP nucleus dengan menghubungkannya melalui faktor hospes hCRM dan dari situ kepada system eksportase seluler nukleus [194]. Akan tetapi NS2/NEP juga terlibat dalam pengendalian replikasi virus, dan studi awal menunjukkan bahwa satu mutasi asam amino tunggal (132T) mengontrol tendensi virus untuk memproduksi partikel pengganggu defektif. [201]. Hanya ada beberapa virus H9N2 yang menunjukkan subtitusi 132V (dari 1000 sekuens yang ada). Tidak ada isolat H9N2 dan H7N9 mengandung treonin pada posisi 32 dari NS/NEP. Baru-baru ini telah ditunjukkan bahwa mutasi M16 sangat berpengaruh dalam mempercepat aktivitas polimerasi virus HPAI H5N1 pada sel manusia tergantung konsentrasinya. NS2/NEP M16 dan 132 tersimpan dengan dalam virus H9N2 dan H7N9.
PB1-F2 adalah produk translasi alternatif dari segmen virus PB1 [32, 35, 75, 179]. Sebelumnya dikarakterisasikan sebagai faktor virulensi pro-apoptotik mitokondrial. Keseluruhan panjang open reading frame PB1-F2 adalah 87-90 asam amino, dan sangat dilestarikan dalam virus AI. Ia juga telah ditemukan selalu ada pada semua isolate pandemi virus influenza yang dikumpulkan selama abad ke 20, akan tetapi tampaknya secara evolusi semakin lama semakin hilang seiring beradaptasinya virus baru pada manusia. PB1-F2 dengan open reading frame sepanjang penuh diperlukan untuk virus HPAI H5N1 agar dapat menimbulkan virulensi penuh pada itik [229]. Pada mamalia, variasi sekuens pada PB1-F2, terutama substitusi N665, memodulasi pathogenesis virus dalam konteks virus flu spanyol H1N1 1918 dan virus HPAI H5N1 [43, 44]. Menariknya, dihapusnya PB1-F2 open reading frame dari virus HPAI H5N1 berakibat meningkatnya virulensi pada ayam [139, 140]. Mutasi tambahan pada PB1-F2 telah diidentifikasi dapat memodulasi virulensi, akan tetapi harus dicatat bahwa aktivitas PB1-F2 tergantung pada konteks dan strainnya [174, 178, 180, 210, 211]. Protein PB1-F2 H9N2 dan H7N9 sebagian bsarnya mengandung 87-90 asam amino. Meskipun begitu penghapusan juga teramati bersama sekuens polimorfisme yang signifikan, meliputi banyak strain yang mengkode susbtitusi N66S. Masih menunggu untuk diketahui apakah penghapusan dan polimorfisme semacam itu memainkan peranan dalam virulensi dan transmisi virus pada unggas dan mamalia.
Sebagai kesimpulan, sifat epistatic dari perubahan molekuler memerlukan penambangan informasi genetik dan fenotipik yang terus menerus dalam rangka mendefinisikan secara lebih baik fitur-fitur yang membawa transmisi virus influenza antar spesies [171]. Di dalam permasalahan ini, virus-virs H9N2 dan H7N9 telah mendapatkan mutasi yang diprediksi mampu meningkatkan kisaran hospes dan potensi pandemiknya.
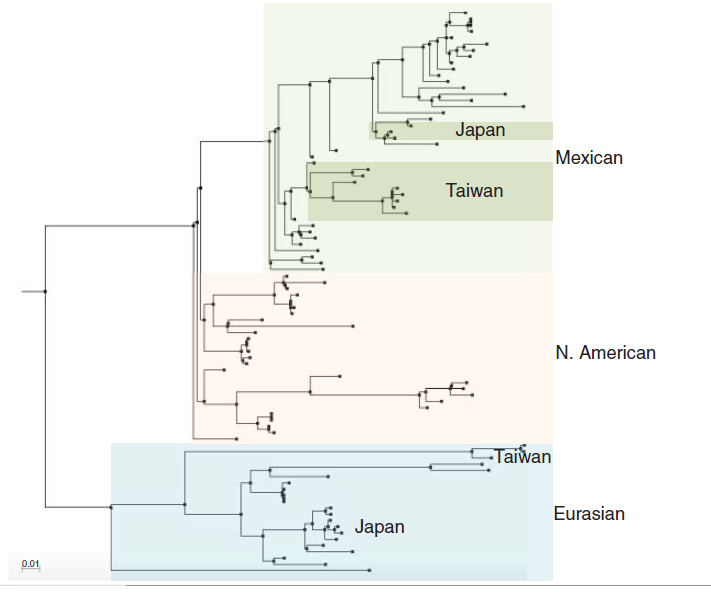
Industri perunggasan di Meksiko menduduki peringkat ke-empat sebagai produser daging ayam terbesar dan peringkat ke-tujuh produser telur di dunia. Pada Maret 1994, virus LPAI H5N2 terdeteksi dalam operasi unggas komersial [280]. Virus ini secara filogeni berhubungan dengan virus-virus H5 lainnya yang berasal dari keturunan Amerika Utara, akan tetapi berbeda dengan virus H5N2 yang menjadi penyebab wabah HPAI di Pennsylvania, Amerika Serikat pada tahun 1983-1984 (Gambar 11.3) [103]. Tingkat penyebaran virus H5N2 kemudian dianalisa dengan serologi, yang menunjukkan bahwa flok-flok positif tanpa gejala yang jelas, baik dari peternakan komersial maupun dari populasi unggas yang dipelihara di belakang rumah, mencakup sebagian wilayah negara Amerika Serikat. Periode istirahat dan program repopulasi dilakukan dengan meliputi kegiatan pengosongan kandang, pembersihan dan desinfeksi fasilitas peternakan. Namun, pada November 1994 virus telah bermutasi menjadi bentuk virus HPAI yang berdampak pada 2 juta ayam petelur di negara bagian Puebla, sebuah wilayah dengan kepadatan unggas tinggi mencakup 25 juta ayam petelur yang tersebar pada lebih dari 100 peternakan. Pada bulan Januari 1995, virus yang serupa muncul di negara bagian Queretaro, berdampak pada 20 juta ayam broiler dan 400.000 peternak. Analisis sekuensing mengungkap adanya mutasi pada tempat pembelahan segmen HA, yaitu dari bentuk sekuens PQRWTR/G yang avirulen menjadi bentuk sekuens PQRKRKTR/G yang bersifat virulen [73, 103]. Kampanye pembasmian dilakukan dengan melibatkan depopulasi peternakan-peternakan terdampak. Pemberishan dan desinfeksi fasilitas peternakan, periode istirahat, penggunaan unggas sentinel sebelum repopulasi dan pengaturan pergerakan unggas dan produk perunggasan yang melalui perbatasan negara bagian. Vaksinasi seluruh unggas juga dilakukan untuk menanggulangi penyebaran penyakit. Keputusan untuk mengimplementasikan program vaksinasi di Meksiko didasarkan pada kesadaran bahwa peternakan-peternakan yang terinfeksi berada pada wilayah dengan kepadatan produksi yang tinggi, yang menyebabkan sulitnya diagnose dan pemusnahan unggas secara cepat. Tambahan lagi, unggas yang diproduksi hanya untuk konsumsi domestik, bukan untuk ekspor. Antara Januari dan Desember tahun 1995, sekitar 380 juga dosis vaksin diberikan kepada populasi unggas terpapar maupun populasi unggas dalam resiko, mencakup sekitar 55% dari keseluruhan populasi unggas di negara Meksiko dan 70% dari keseluruhan unggas peternakan komersial. Vaksinasi terhadap ayam petelur dengan siklus yang Panjang dilakukan di negara-negara bagian dimana kasus-kasus LPAI telah terdeteksi. Bulan Mei 1995 menandai berakhirnya isolasi terakhir HPAI H5N2 di Meksiko, dan pada Desember 1995 pemerintah Meksiko telag menyatakan bahwa negara mereka telah bebas dari virus ini, meskipun virus LPAI H5N2 masih terus bersirkulasi. Tidak ada kasus pada manusia yang dilaporkan, dan tidak ada bukti adanya infeksi pada manusia. Perkiraan biaya program tanggap darurat adalah 49 juta USD, mencakup operasional, biosekuriti, vaksinasi dan nilai unggas yang mati dan dimusnahkan [1]. Sejak Juni 1995, virus LPAI H5N2 tetap terus bersirkulasi di Meksiko dan telah menyebar pada unggas di Guatemala dan El Salvador. Bukti adanya antigenic drift yang dihubungkan dengan penggunaan vaksin, kemungkinan dari vaksinasi yang tidak tepat maupun tidak sesuai, telah terdeteksi, yang mungkin saha merupakan efek yang tidak diinginkan dari produksi strain yang memiliki kemampuan beralih hospes [133]. Juga penting untuk diingat bahwa strain H5N2 yang berasal dari Meksiko, yang mungkin diperuntukkan untuk dipakai sebagai vaksin, telah menyebabkan dua wabah terpisah di Jepang dan Taiwan (Gambar 11.3) [39, 131, 204, 205]. Di Taiwan, virus H5N2 muncul sebagai reasortan yang membawa segmen gen permukaan dari virus H5N2 asal Meksiko dan segmen gen internal dari virus H6N1 asal Eurasia yang telah bersirkulasi lebih dulu di negara tersebut. Reasortan virus H5N2 terus saja bersirkulasi di Taiwan, menambahkan pada keberagaman virus LPAI di wilayah tersebut.
Gambar 11.3 Pohon filogeni H5 HA menunjukkan hubungan filogeni besar antara keturunan virus-virus H5N2 dari Amerika Utara (warna oranye muda) dan Eurasia (biru muda). Analisis dan penyesuaian filogeni telah dilakukan seperti yang telah dideskripsikan pada Gambar 11.1. Virus-virus Meksiko (ditunjukkan dengan warna hijau muda) membentuk sebuah garis evolusi independent yang muncul dari nenek moyang dari garis Amerika Utara. Virus-virus H5N2 Meksiko yang diturunkan dari vaksin telah diisolasi dari wabah-wabah LPAI di Jepang dan Taiwan. Di Taiwan, segmen gen permukaan virus H5N2 yang diturunkan dari Meksiko telah mengalami reasortasi dengan virus LPAI Taiwan, dan status endeminya masih belum diketahui. Lihat bagian Plate untuk melihat representasi warna-warna yang disajikan pada gambar ini.
Seiring meluasnya perkembangan produksi perunggasan, meningkat pula kemungkinan bahwa pathogen dari unggas akan mendapat kesempatan berpindah ke hospes lain. Potensi zoonosis dari virus LPAI juga telah diketahui dengan baik. Selama kurun waktu 25 tahun terakhir ini telah terlihat meningkatnya jumlah virus LPAI dan (HPAI) yang telah menetap menjadi endemic dalam populasi unggas, dan bersaam dengan itu pula telah terlihat persebaran pada wilayah-wilayah geografis yang belum pernah terjadi sebelumnya. Kombinasi faktor-faktor agricultural, environmental, sosisal dan ekonomi telah membawa pada munculnya virus LPAI yang memiliki potensi pandemic. Di masa depan diperlukan upaya-upaya dalam menyelaraskan sistem produksi yang berkelanjutan, praktek-praktek kultural dalam beternak, serta startegi-stategi intervensi baik tradisional maupun yang sama sekali baru untuk dapat menurunkan beban yang disebabkan penyakit LPAI (dan pathogen-patogen unggas lainnya), yang pada akhirnya akan meminimalisir resiko zoonosis dan kesehatan masyarakat.
Abdel-Moneim, A. S., M. A. Afifi and M. F. El-Kady, 2012: Isolation and mutation trend analysis of influenza A virus subtype H9N2 in Egypt. Virology Journal,9, 1-9.
Abdelwhab, E. M., J. Veits and T. C. Mettenleiter, 2014: Prevalence and control of H7 avian influenza viruses in birds and humans. Epidemiology & Infection,142, 896-920.
Abolnik, C., S. Bisschop, T. Gerdes, A. Olivier and R. Horner, 2007: Outbreaks of avian influenza H6N2 viruses in chickens arose by a reassortment of H6N8 and H9N2 ostrich viruses. Virus Genes,34, 37-45.
Abolnik, C., S. P. R. Bisschop, G. H. Gerdes, A. J. Olivier and R. F. Horner, 2007: Phylogenetic analysis of low-pathogenicity avian influenza H6N2 viruses from chicken outbreaks (2001–2005) suggest that they are reassortants of historic ostrich low-pathogenicity avian influenza H9N2 and H6N8 viruses. Avian Diseases,51, 279-284.
Akey, B. L., 2003: Low-pathogenicity H7N2 avian influenza outbreak in Virginia during 2002. Avian Diseases,47, 1099-1103.
Alexander, D. J., 2000: A review of avian influenza in different bird species. Veterinary Microbiology,74, 3-13.
Amirsalehy, H., H. Nili and A. Mohammadi, 2012: Can dogs carry the global pandemic candidate avian influenza virus H9N2? Australian Veterinary Journal,90, 341-345.
Arafa, A.-S., N. Hagag, A. Erfan, W. Mady, M. El-Husseiny, A. Adel and S. Nasef, 2012: Complete genome characterization of avian influenza virus subtype H9N2 from a commercial quail flock in Egypt. Virus Genes,45, 283-294.
Arafa, A.-S., N. M. Hagag, N. Yehia, A. M. Zanaty, M. M. Naguib and S. A. Nasef, 2012: Effect of cocirculation of highly pathogenic avian influenza H5N1 subtype with low pathogenic H9N2 subtype on the spread of infections. Avian Diseases,56, 849-857.
Ayllon, J., P. Domingues, R. Rajsbaum, L. Miorin, M. Schmolke, B. G. Hale and A. García-Sastre, 2014: A single amino acid substitution in the novel H7N9 influenza A virus NS1 protein increases CPSF30 binding and virulence. Journal of Virology,88, 12146-12151.
Ayllon, J. and A. García-Sastre, 2014: The NS1 protein: a multitasking virulence factor. Current Topics in Microbiology and Immunology,386, 73-107.
Bahl, A. K., B. S. Pomeroy and B. C. Easterday, 1975: Isolation of two turkey influenza A viruses in Minnesota. Avian Diseases,19, 374-378.
Banet-Noach, C., S. Perk, L. Simanov, N. Grebenyuk, E. Rozenblut, S. Pokamunski, M. Pirak, Y. Tendler and A. Panshin, 2007: H9N2 influenza viruses from Israeli poultry: a five-year outbreak. Avian Diseases,51, 290-296.
Banks, J., E. S. Speidel, E. Moore, L. Plowright, A. Piccirillo, I. Capua, P. Cordioli, A. Fioretti and D. J. Alexander, 2001: Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Archives of Virology,146, 963-973.
Barbour, E. K., V. K. Sagherian, N. K. Sagherian, S. K. Dankar, L. S. Jaber, N. N. Usayran and M. T. Farran, 2006: Avian influenza outbreak in poultry in the Lebanon and transmission to neighbouring farmers and swine. Veterinaria Italiana,42, 77-85.
Baron, J., C. Tarnow, D. Mayoli-Nüssle, E. Schilling, D. Meyer, M. Hammami, F. Schwalm, T. Steinmetzer, Y. Guan and W. Garten, 2013: Matriptase, HAT, and TMPRSS2 activate the hemagglutinin of H9N2 influenza A viruses. Journal of Virology,87, 1811-1820.
Belser, J. A., C. T. Davis, A. Balish, L. E. Edwards, H. Zeng, T. R. Maines, K. M. Gustin, I. L. Martínez, R. Fasce and N. J. Cox, 2013: Pathogenesis, transmissibility, and ocular tropism of a highly pathogenic avian influenza A (H7N3) virus associated with human conjunctivitis. Journal of virology,87, 5746-5754.
Belser, J. A., K. M. Gustin, M. B. Pearce, T. R. Maines, H. Zeng, C. Pappas, X. Sun, P. J. Carney, J. M. Villanueva and J. Stevens, 2013: Pathogenesis and transmission of avian influenza A (H7N9) virus in ferrets and mice. Nature,501, 556-559.
Belser, J. A., X. Lu, T. R. Maines, C. Smith, Y. Li, R. O. Donis, J. M. Katz and T. M. Tumpey, 2007: Pathogenesis of avian influenza (H7) virus infection in mice and ferrets: enhanced virulence of Eurasian H7N7 viruses isolated from humans. Journal of Virology,81, 11139-11147.
Belser, J. A. and T. M. Tumpey, 2014: Mammalian models for the study of H7 virus pathogenesis and transmission. Current Topics in Microbiology and Immunology,385, 275-305.
Bi, Y., L. Lu, J. Li, Y. Yin, Y. Zhang, H. Gao, Z. Qin, B. Zeshan, J. Liu and L. Sun, 2011: Novel genetic reassortants in H9N2 influenza A viruses and their diverse pathogenicity to mice. Virology Journal,8, 1-11.
Blair, P. J., S. D. Putnam, W. S. Krueger, C. Chum, T. F. Wierzba, G. L. Heil, C. Y. Yasuda, M. Williams, M. R. Kasper and J. A. Friary, 2013: Evidence for avian H9N2 influenza virus infections among rural villagers in Cambodia. Journal of Infection and Public Health,6, 69-79.
Brown, I. H., J. Banks, R. J. Manvell, S. C. Essen, W. Shell, M. Slomka, B. Londt and D. J. Alexander, 2006: Recent epidemiology and ecology of influenza A viruses in avian species in Europe and the Middle East. Developments in Biologicals,124, 45-50.
Bulaga, L. L., L. Garber, D. A. Senne, T. J. Myers, R. Good, S. Wainwright, S. Trock and D. L. Suarez, 2003: Epidemiologic and surveillance studies on avian influenza in live-bird markets in New York and New Jersey, 2001. Avian Diseases,47, 996-1001.
Bussey, K. A., E. A. Desmet, J. L. Mattiacio, A. Hamilton, B. Bradel-Tretheway, H. E. Bussey, B. Kim, S. Dewhurst and T. Takimoto, 2011: PA residues in the 2009 H1N1 pandemic influenza virus enhance avian influenza virus polymerase activity in mammalian cells. Journal of Virology,85, 7020-7028.
Butt, A. M., S. Siddique, M. Idrees and Y. Tong, 2010: Avian influenza A (H9N2): computational molecular analysis and phylogenetic characterization of viral surface proteins isolated between 1997 and 2009 from the human population. Virology Journal,7, 1-11.
Butt, K. M., G. J. D. Smith, H. Chen, L. J. Zhang, Y. H. C. Leung, K. M. Xu, W. Lim, R. G. Webster, K. Y. Yuen and J. S. M. Peiris, 2005: Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. Journal of Clinical Microbiology 43, 5760-5767.
Centers for Disease Control and Prevention (CDC). 2013. Emergence of avian influenza A (H7N9) virus causing severe human illness—China, February–April 2013. Morbidity and Mortality Weekly Report,62, 366-371.
Centers for Disease Control and Prevention (CDC). 2012. Notes from the field: Highly pathogenic avian influenza A (H7N3) virus infection in two poultry workers–Jalisco, Mexico, July 2012. Morbidity and Mortality Weekly Report,61, 726-727.
Chang, F. Y., T. Y. Lin and W. T. Chiu, 2013: Human infection with H7N9 virus. New England Journal of Medicine,369, 879-880.
Chang, S. Y., P. H. Lin, J. C. Tsai, C. C. Hung and S. C. Chang, 2013: The first case of H7N9 influenza in Taiwan. The Lancet,381,
Chanturiya, A. N., G. Basanez, U. Schubert, P. Henklein, J. W. Yewdell and J. Zimmerberg, 2004: PB1-F2, an influenza A virus-encoded proapoptotic mitochondrial protein, creates variably sized pores in planar lipid membranes. Journal of Virology,78, 6304-6312.
Chen, E., Y. Chen, L. Fu, Z. Chen, Z. Gong, H. Mao, D. Wang, M. Y. Ni, P. Wu and Z. Yu, 2013: Human infection with avian influenza A (H7N9) virus re-emerges in China in winter 2013. Euro Surveillance,18,
Chen, H., H. Yuan, R. Gao, J. Zhang, D. Wang, Y. Xiong, G. Fan, F. Yang, X. Li and J. Zhou, 2014: Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: a descriptive study. The Lancet,383, 714-721.
Chen, W., P. A. Calvo, D. Malide, J. Gibbs, U. Schubert, I. Bacik, S. Basta, R. O’Neill, J. Schickli and P. Palese, 2001: A novel influenza A virus mitochondrial protein that induces cell death. Nature Medicine,7, 1306-1312.
Chen, X., Z. Yang, Y. Lu, Q. Xu, Q. Wang and L. Chen, 2013: Clinical features and factors associated with outcomes of patients infected with a Novel Influenza A (H7N9) virus: a preliminary study. Plos One,8,
Cheng, H. W. B., C. W. Li, K. Y. Chan and M. K. Sham, 2014: The first confirmed case of human avian influenza A (H7N9) in Hong Kong and the suspension of volunteer services: impact on palliative care. Journal of Pain and Symptom Management,47, e5-7.
Cheng, K., Z. Yu, H. Chai, W. Sun, Y. Xin, Q. Zhang, J. Huang, K. Zhang, X. Li and S. Yang, 2014: PB2-E627K and PA-T97I substitutions enhance polymerase activity and confer a virulent phenotype to an H6N1 avian influenza virus in mice. Virology,468, 207-213.
Cheng, M. C., K. Soda, M.-S. Lee, S. H. Lee, Y. Sakoda, H. Kida and C. H. Wang, 2010: Isolation and characterization of potentially pathogenic H5N2 influenza virus from a chicken in Taiwan in 2008. Avian Diseases,54, 885-893.
Cheng, V. C. C., J. F. W. Chan, X. Wen, W. L. Wu, T. L. Que, H. Chen, K. H. Chan and K. Y. Yuen, 2011: Infection of immunocompromised patients by avian H9N2 influenza A virus. Journal of Infection,62, 394-399.
Choi, Y. K., H. Ozaki, R. J. Webby, R. G. Webster, J. S. Peiris, L. Poon, C. Butt, Y. H. C. Leung and Y. Guan, 2004: Continuing evolution of H9N2 influenza viruses in Southeastern China. Journal of virology,78, 8609-8614.
Chu, Y. C., C. L. Cheung, C. Y. H. Leung, L. L. M. Poon, H. Chen, J. S. M. Peiris and Y. Guan, 2011: Continuing evolution of H9N2 influenza viruses endemic in poultry in southern China. Influenza and Other Respiratory Viruses,5 (Suppl. 1), 68-71.
Conenello, G. M., J. R. Tisoncik, E. Rosenzweig, Z. T. Varga, P. Palese and M. G. Katze, 2011: A single N66S mutation in the PB1-F2 protein of influenza A virus increases virulence by inhibiting the early interferon response in vivo. Journal of Virology,85, 652-662.
Conenello, G. M., D. Zamarin, L. A. Perrone, T. Tumpey and P. Palese, 2007: A single mutation in the PB1-F2 of H5N1 (HK/97) and 1918 influenza A viruses contributes to increased virulence. Plos One,3,
Cong, Y. L., J. Pu, Q. F. Liu, S. Wang, G. Z. Zhang, X. L. Zhang, W. X. Fan, E. G. Brown and J. H. Liu, 2007: Antigenic and genetic characterization of H9N2 swine influenza viruses in China. Journal of General Virology,88, 2035-2041.
Cong, Y. L., C. F. Wang, C. M. Yan, J. S. Peng, Z. L. Jiang and J. H. Liu, 2008: Swine infection with H9N2 influenza viruses in China in 2004. Virus Genes,36, 461-469.
Dalby, A. R. and M. Iqbal, 2014: A global phylogenetic analysis in order to determine the host species and geography dependent features present in the evolution of avian H9N2 influenza hemagglutinin. PeerJ,2,
Davidson, I., A. Fusaro, A. Heidari, I. Monne and G. Cattoli, 2014: Molecular evolution of H9N2 avian influenza viruses in Israel. Virus Genes,48, 457-463.
Davidson, I., I. Shkoda, N. Golender, S. Perk, K. Lapin, Y. Khinich and A. Panshin, 2013: Genetic characterization of HA gene of low pathogenic H9N2 influenza viruses isolated in Israel during 2006–2012 periods. Virus Genes,46, 255-263.
Davison, S., R. J. Eckroade and A. F. Ziegler, 2003: A review of the 1996–98 nonpathogenic H7N2 avian influenza outbreak in Pennsylvania. Avian Diseases,47, 823-827.
Dong, G., J. Luo, H. Zhang, C. Wang, M. Duan, T. J. Deliberto, D. L. Nolte, G. Ji and H. He, 2011: Phylogenetic diversity and genotypical complexity of H9N2 influenza A viruses revealed by genomic sequence analysis. PloS One,6,
Dong, W., X. Li Feng, W. Cun Lian, X. Ming Ju, Z. Rui Hua, L. Ying and X. Tong, 2012: A mouse model of swine influenza virus H9N2 infection with acute lung injury. Acta Virologica,56, 227-233.
Dortmans, J. C. F. M., J. Dekkers, I. N. A. Wickramasinghe, M. H. Verheije, P. J. M. Rottier, F. J. M. Van Kuppeveld, E. De Vries and C. A. M. De Haan, 2013: Adaptation of novel H7N9 influenza A virus to human receptors. Scientific reports,3, 1-7.
van Beest Holle, M. D. R., A. Meijer, M. Koopmans and C. M. De Jager, 2005: Human-to-human transmission of avian influenza A/H7N7, The Netherlands, 2003. Euro Surveillance,10, 3-4.
Dundon, W. G. and I. Capua, 2009: A closer look at the NS1 of influenza virus. Viruses,1, 1057-1072.
Eames, K. T. D., C. Webb, K. Thomas, J. Smith, R. Salmon and J. M. F. Temple, 2010: Assessing the role of contact tracing in a suspected H7N2 influenza A outbreak in humans in Wales. BMC Infectious Diseases,10, 1-6.
Easterday, B. C., D. O. Trainer, B. Tůmová and H. G. Pereira, 1968: Evidence of infection with influenza viruses in migratory waterfowl. Nature,219, 523-524.
Easterday, B. C. and B. Tumova, 1972: Avian influenza viruses: in avian species and the natural history of influenza. Advances in Veterinary Science and Comparative Medicine.
Ebrahimi, S. M., S. Ziapour, M. Tebianian, M. Dabaghian and M. Mohammadi, 2011: Study of infection with an Iranian field-isolated H9N2 avian influenza virus in vaccinated and unvaccinated Japanese quail. Avian Diseases,55, 195-200.
Fan, S., G. Deng, J. Song, G. Tian, Y. Suo, Y. Jiang, Y. Guan, Z. Bu, Y. Kawaoka and H. Chen, 2009: Two amino acid residues in the matrix protein M1 contribute to the virulence difference of H5N1 avian influenza viruses in mice. Virology,384, 28-32.
Fan, S., M. Hatta, J. H. Kim, M. Q. Le, G. Neumann and Y. Kawaoka, 2014: Amino acid changes in the influenza A virus PA protein that attenuate avian H5N1 viruses in mammals. Journal of Virology,88, 13737-13746.
Fan, S., C. A. Macken, C. Li, M. Ozawa, H. Goto, N. F. N. Iswahyudi, C. A. Nidom, H. Chen, G. Neumann and Y. Kawaoka, 2013: Synergistic effect of the PDZ and p85β-binding domains of the NS1 protein on virulence of an avian H5N1 influenza A virus. Journal of Virology,87, 4861-4871.
Fang, L. Q., X. L. Li, K. Liu, Y. J. Li, H. W. Yao, S. Liang, Y. Yang, Z. J. Feng, G. C. Gray and W. C. Cao, 2013: Mapping spread and risk of avian influenza A (H7N9) in China. Scientific reports,3, 1-8.
Feng, Y., H. Mao, C. Xu, J. Jiang, Y. Chen, J. Yan, J. Gao, Z. Li, S. Xia and Y. Lu, 2013: Origin and characteristics of internal genes affect infectivity of the novel avian-origin influenza A (H7N9) virus. Plos One,8,
Firth, A. E., B. W. Jagger, H. M. Wise, C. C. Nelson, K. Parsawar, N. M. Wills, S. Napthine, J. K. Taubenberger, P. Digard and J. F. Atkins, 2012: Ribosomal frameshifting used in influenza A virus expression occurs within the sequence UCC_UUU_CGU and is in the+ 1 direction. Open Biology,2,
Fouchier, R. A. M., P. M. Schneeberger, F. W. Rozendaal, J. M. Broekman, S. A. G. Kemink, V. Munster, T. Kuiken, G. F. Rimmelzwaan, M. Schutten and G. J. J. Van Doornum, 2004: Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proceedings of the National Academy of Sciences,101, 1356-1361.
Fournié, G., F. J. Guitian, P. Mangtani and A. C. Ghani, 2011: Impact of the implementation of rest days in live bird markets on the dynamics of H5N1 highly pathogenic avian influenza. Journal of the Royal Society Interface,8, 1079-1089.
Fournié, G. and D. U. Pfeiffer, 2014: Can closure of live poultry markets halt the spread of H7N9? The Lancet,383, 496-497.
Fusaro, A., I. Monne, A. Salviato, V. Valastro, A. Schivo, N. M. Amarin, C. Gonzalez, M. M. Ismail, A.-R. Al-Ankari and M. H. Al-Blowi, 2011: Phylogeography and evolutionary history of reassortant H9N2 viruses with potential human health implications. Journal of Virology,85, 8413-8421.
Gambaryan, A., R. Webster and M. Matrosovich, 2002: Differences between influenza virus receptors on target cells of duck and chicken. Archives of Virology,147, 1197-1208.
Gao, H. N., H. Z. Lu, B. Cao, B. Du, H. Shang, J. H. Gan, S. H. Lu, Y. D. Yang, Q. Fang and Y. Z. Shen, 2013: Clinical findings in 111 cases of influenza A (H7N9) virus infection. New England Journal of Medicine,368, 2277-2285.
Gao, R., B. Cao, Y. Hu, Z. Feng, D. Wang, W. Hu, J. Chen, Z. Jie, H. Qiu and K. Xu, 2013: Human infection with a novel avian-origin influenza A (H7N9) virus. New England Journal of Medicine,368, 1888-1897.
Garcia, M., J. M. Crawford, J. W. Latimer, E. Rivera-Cruz and M. L. Perdue, 1996: Heterogeneity in the haemagglutinin gene and emergence of the highly pathogenic phenotype among recent H5N2 avian influenza viruses from Mexico. Journal of General Virology,77, 1493-1504.
Giannecchini, S., V. Clausi, L. Di Trani, E. Falcone, C. Terregino, A. Toffan, F. Cilloni, M. Matrosovich, A. S. Gambaryan and N. V. Bovin, 2010: Molecular adaptation of an H7N3 wild duck influenza virus following experimental multiple passages in quail and turkey. Virology,408, 167-173.
Gibbs, J. S., D. Malide, F. Hornung, J. R. Bennink and J. W. Yewdell, 2003: The influenza A virus PB1-F2 protein targets the inner mitochondrial membrane via a predicted basic amphipathic helix that disrupts mitochondrial function. Journal of Virology,77, 7214-7224.
Golebiewski, L., H. Liu, R. T. Javier and A. P. Rice, 2011: The avian influenza virus NS1 ESEV PDZ binding motif associates with Dlg1 and Scribble to disrupt cellular tight junctions. Journal of Virology,85, 10639-10648.
Gonzales, J. L., J. A. Stegeman, G. Koch, S. J. de Wit and A. R. W. Elbers, 2013: Rate of introduction of a low pathogenic avian influenza virus infection in different poultry production sectors in the Netherlands. Influenza and Other Respiratory Viruses,7, 6-10.
González, S., T. Zürcher and J. Ortín, 1996: Identification of two separate domains in the influenza virus PB1 protein involved in the interaction with the PB2 and PA subunits: a model for the viral RNA polymerase structure. Nucleic Acids Research,24, 4456-4463.
Gou, Y., J. Xie and M. Wang, 2000: A strain of influenza A H9N2 virus repeatedly isolated from human population in China. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi,14, 209-212.
Gu, H. X., S. L. Hu and Z. G. Yang, 1996: The Relationship between Integrated Fish Farming and Human Influenza Pandemic. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi,17, 29-32.
Gu, H. X., Z. G. Yang and X. Y. Jin, 1996: An epidemiological study on influenza pandemic. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi,17, 77-79.
Guan, Y., K. F. Shortridge, S. Krauss and R. G. Webster, 1999: Molecular characterization of H9N2 influenza viruses: were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proceedings of the National Academy of Sciences,96, 9363-9367.
Guo, C. T., N. Takahashi, H. Yagi, K. Kato, T. Takahashi, S. Q. Yi, Y. Chen, T. Ito, K. Otsuki and H. Kida, 2007: The quail and chicken intestine have sialyl-galactose sugar chains responsible for the binding of influenza A viruses to human type receptors. Glycobiology,17, 713-724.
Guo, Y., J. Li and X. Cheng, 1999: Discovery of men infected by avian influenza A (H9N2) virus. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi,13, 105-108.
Guo, Y. J., S. Krauss, D. A. Senne, I. P. Mo, K. S. Lo, X. P. Xiong, M. Norwood, K. F. Shortridge, R. G. Webster and Y. Guan, 2000: Characterization of the pathogenicity of members of the newly established H9N2 influenza virus lineages in Asia. Virology,267, 279-288.
Hai, R., M. Schmolke, V. H. Leyva-Grado, R. R. Thangavel, I. Margine, E. L. Jaffe, F. Krammer, A. Solórzano, A. García-Sastre and P. J. N. c. Palese, 2013: Influenza A (H7N9) virus gains neuraminidase inhibitor resistance without loss of in vivo virulence or transmissibility. Nature Communications,4, 1-9.
Hale, B. G., 2014: Conformational plasticity of the influenza A virus NS1 protein. Journal of General Virology,95, 2099-2105.
Hale, B. G., R. E. Randall, J. Ortín and D. Jackson, 2008: The multifunctional NS1 protein of influenza A viruses. Journal of General Virology,89, 2359-2376.
Halvorson, D. 2008. Control of low pathogenicity avian influenza in USA. In: Avian Influenza, D. E. Swayne, ed. Blackwell Publishing: Ames, IA. 513-536.
Halvorson, D., D. D. Frame, A. J. Frinedshuh, and D. P. Shaw. 1997. Outbreaks of low pathogenicity avian influenza in USA. In: Proceedings of The Fourth International Symposium on Avian Influenza, D. Swayne and R. D. Slemons, eds. United States Animal Health Association: Tallahassee, FL. 36-46.
Halvorson, D., D. Karunakaran, D. Senne, C. Kelleher, C. Bailey, A. Abraham, V. Hinshaw and J. Newman, 1983: Epizootiology of avian influenza: simultaneous monitoring of sentinel ducks and turkeys in Minnesota. Avian Diseases,27, 77-85.
Harder, T. and C. Grund, 2014: Ausbreitung und Einddammungmöglichkeiten von H9N2 Infektionen in Geflugel. 87th Fachgesprach Uber Geflugelkrankheiten. 47-48.
Hatta, M., P. Gao, P. Halfmann and Y. Kawaoka, 2001: Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science,293, 1840-1842.
Hatta, M., Y. Hatta, J. H. Kim, S. Watanabe, K. Shinya, T. Nguyen, P. S. Lien, Q. M. Le and Y. Kawaoka, 2007: Growth of H5N1 influenza A viruses in the upper respiratory tracts of mice. PLoS Pathogens,3, 1374-1379.
Hayward, J. J., E. J. Dubovi, J. M. Scarlett, S. Janeczko, E. C. Holmes and C. R. Parrish, 2010: Microevolution of canine influenza virus in shelters and its molecular epidemiology in the United States. Journal of Virology,84, 12636-12645.
He, X., J. Zhou, M. Bartlam, R. Zhang, J. Ma, Z. Lou, X. Li, J. Li, A. Joachimiak and Z. Zeng, 2008: Crystal structure of the polymerase PA C–PB1 N complex from an avian influenza H5N1 virus. Nature,454, 1123-1126.
He, Y., P. Liu, S. Tang, Y. Chen, E. Pei, B. Zhao, H. Ren, J. Li, Y. Zhu and H. Zhao, 2014: Live poultry market closure and control of avian influenza A (H7N9), Shanghai, China. Emerging Infectious Diseases,20,
Herfst, S., E. J. Schrauwen, M. Linster, S. Chutinimitkul, E. de Wit, V. J. Munster, E. M. Sorrell, T. M. Bestebroer, D. F. Burke and D. J. Smith, 2012: Airborne transmission of influenza A/H5N1 virus between ferrets. Science,336, 1534-1541.
Homme, P. J. and B. C. Easterday, 1970: Avian influenza virus infections. III. Antibody response by turkeys to influenza A/turkey/Wisconsin/1966 virus. Avian Diseases,14, 277-284.
Homme, P. J. and B. C. Easterday, 1970: Avian influenza virus infections. III. Antibody response by turkeys to influenza A/turkey/Wisconsin/1966 virus. Avian Diseases,14, 277-284.
Homme, P. J. and B. C. Easterday, 1970: Avian influenza virus infections. IV. Response of pheasants, ducks, and geese to Influenza A/Turkey/Wisconsin/1966 virus. Avian Diseases,14, 285-290.
Homme, P. J., B. C. Easterday and D. P. Anderson, 1970: Avian influenza virus infections. II. Experimental epizootiology of influenza A/Turkey/Wisconsin/1966 virus in turkeys. Avian Diseases,14, 240-247.
Horimoto, T., E. Rivera, J. Pearson, D. Senne, S. Krauss, Y. Kawaoka and R. G. Webster, 1995: Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexico. Virology,213, 223-230.
Hossain, M. J., D. Hickman and D. R. Perez, 2008: Evidence of expanded host range and mammalian-associated genetic changes in a duck H9N2 influenza virus following adaptation in quail and chickens. PloS One,3,
Hu, W., W. Zhang, X. Huang, A. Clements, K. Mengersen and S. Tong, 2015: Weather variability and influenza A (H7N9) transmission in Shanghai, China: A Bayesian spatial analysis. Environmental Research,136, 405-412.
Hu, Y., S. Lu, Z. Song, W. Wang, P. Hao, J. Li, X. Zhang, H.-L. Yen, B. Shi and T. Li, 2013: Association between adverse clinical outcome in human disease caused by novel influenza A H7N9 virus and sustained viral shedding and emergence of antiviral resistance. The Lancet,381, 2273-2279.
Huang, R., J. Liu, W. Liang, A. Wang, Z. Liu, Y. Yang, J. Lv, Y. Bao, Y. Gao and Z. Miao, 2014: Response profiles of cytokines and chemokines against avian H9N2 influenza virus within the mouse lung. Medical Microbiology and Immunology,203, 109-114.
Huang, R., A. R. Wang, Z. H. Liu, W. Liang, X. X. Li, Y. J. Tang, Z. M. Miao and T. J. Chai, 2013: Seroprevalence of avian influenza H9N2 among poultry workers in Shandong Province, China. European Journal of Clinical Microbiology and Infectious Diseases,32, 1347-1351.
Huang, Z., D. Fang, P. Lv, X. Bian, X. Ruan, Y. Yan and J. Zhou, 2012: Differential cellular immune responses between chickens and ducks to H9N2 avian influenza virus infection. Veterinary Immunology and Immunopathology,150, 169-180.
Ilyushina, N. A., M. F. Ducatez, J. E. Rehg, B. M. Marathe, H. Marjuki, N. V. Bovin, R. G. Webster and R. J. Webby, 2010: Does pandemic A/H1N1 virus have the potential to become more pathogenic? MBio,1, e00249-00210.
Ilyushina, N. A., A. M. Khalenkov, J. P. Seiler, H. L. Forrest, N. V. Bovin, H. Marjuki, S. Barman, R. G. Webster and R. J. Webby, 2010: Adaptation of pandemic H1N1 influenza viruses in mice. Journal of Virology,84, 8607-8616.
Jackson, D., M. J. Hossain, D. Hickman, D. R. Perez and R. A. Lamb, 2008: A new influenza virus virulence determinant: the NS1 protein four C-terminal residues modulate pathogenicity. Proceedings of the National Academy of Sciences,105, 4381-4386.
Jagger, B. W., H. M. Wise, J. C. Kash, K. A. Walters, N. M. Wills, Y. L. Xiao, R. L. Dunfee, L. M. Schwartzman, A. Ozinsky and G. L. Bell, 2012: An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science,337, 199-204.
Jakhesara, S. J., V. D. Bhatt, N. V. Patel, K. S. Prajapati and C. G. Joshi, 2014: Isolation and characterization of H9N2 influenza virus isolates from poultry respiratory disease outbreak. Springer Plus,3, 1-8.
Kalthoff, D., J. Bogs, C. Grund, K. Tauscher, J. P. Teifke, E. Starick, T. Harder and M. Beer, 2014: Avian influenza H7N9/13 and H7N7/13: a comparative virulence study in chickens, pigeons, and ferrets. Journal of Virology,88, 9153-9165.
Ke, C., J. Lu, J. Wu, D. Guan, L. Zou, T. Song, L. Yi, X. Zeng, L. Liang and H. Ni, 2014: Circulation of reassortant influenza A (H7N9) viruses in poultry and humans, Guangdong Province, China, 2013. Emerging Infectious Diseases,20,
Ke, Y., Y. Wang, S. Liu, J. Guo, W. Zhang, X. Yuan, N. Zhang, Z. Wang, H. Song and L. Huang, 2013: High severity and fatality of human infections with avian influenza A (H7N9) infection in China. Clinical Infectious Diseases,57, 1506-1507.
Kim, J. A., S. H. Cho, H. S. Kim and S. H. Seo, 2006: H9N2 influenza viruses isolated from poultry in Korean live bird markets continuously evolve and cause the severe clinical signs in layers. Veterinary Microbiology,118, 169-176.
Kimble, B., G. R. Nieto and D. R. Perez, 2010: Characterization of influenza virus sialic acid receptors in minor poultry species. Virology Journal,7, 1-10.
Kimble, J. B., M. Angel, H. Wan, T. C. Sutton, C. Finch and D. R. Perez, 2014: Alternative reassortment events leading to transmissible H9N1 influenza viruses in the ferret model. Journal of Virology,88, 66-71.
Kimble, J. B., E. Sorrell, H. Shao, P. L. Martin and D. R. Perez, 2011: Compatibility of H9N2 avian influenza surface genes and 2009 pandemic H1N1 internal genes for transmission in the ferret model. Proceedings of the National Academy of Sciences,108, 12084-12088.
Koopmans, M., B. Wilbrink, M. Conyn, G. Natrop, H. van der Nat, H. Vennema, A. Meijer, J. van Steenbergen, R. Fouchier and A. Osterhaus, 2004: Transmission of H7N7 avian influenza A virus to human beings during a large outbreak in commercial poultry farms in the Netherlands. The Lancet,363, 587-593.
Krug, R. M., W. Yuan, D. L. Noah and A. G. Latham, 2003: Intracellular warfare between human influenza viruses and human cells: the roles of the viral NS1 protein. Virology,309, 181-189.
Kumar, M., H. Liu and A. P. Rice, 2012: Regulation of interferon-β by MAGI-1 and its interaction with influenza A virus NS1 protein with ESEV PBM. Plos One,7,
Kuo, R.-L., C. Zhao, M. Malur and R. M. Krug, 2010: Influenza A virus strains that circulate in humans differ in the ability of their NS1 proteins to block the activation of IRF3 and interferon-β transcription. Virology,408, 146-158.
Kwon, J. S., H. J. Lee, D. H. Lee, Y. J. Lee, I. P. Mo, S. S. Nahm, M. J. Kim, J. B. Lee, S. Y. Park and I. S. Choi, 2008: Immune responses and pathogenesis in immunocompromised chickens in response to infection with the H9N2 low pathogenic avian influenza virus. Virus Research,133, 187-194.
Lam, T. T. Y., J. Wang, Y. Shen, B. Zhou, L. Duan, C. L. Cheung, C. Ma, S. J. Lycett, C. Y. H. Leung and X. Chen, 2013: The genesis and source of the H7N9 influenza viruses causing human infections in China. Nature,502, 241-244.
Lavoie, E. T., E. M. Sorrell, D. R. Perez and M. A. Ottinger, 2007: Immunosenescence and age-related susceptibility to influenza virus in Japanese quail. Developmental and Comparative Immunology,31, 407-414.
Le, Q. M., Y. Sakai-Tagawa, M. Ozawa, M. Ito and Y. Kawaoka, 2009: Selection of H5N1 influenza virus PB2 during replication in humans. Journal of Virology,83, 5278-5281.
Lee, C., D. Song, B. Kang, D. Kang, J. Yoo, K. Jung, G. Na, K. Lee, B. Park and J. Oh, 2009: A serological survey of avian origin canine H3N2 influenza virus in dogs in Korea. Veterinary Microbiology,137, 359-362
Lee, C.-C. D., H. Zhu, P. Y. Huang, L. Peng, Y. C. Chang, C. H. Yip, Y. T. Li, C. L. Cheung, R. Compans and C. Yang, 2014: Emergence and evolution of avian H5N2 influenza viruses in chickens in Taiwan. Journal of Virology,88, 5677-5686.
Lee, C. H., S. H. Byun, Y. J. Lee and I. P. Mo, 2012: Genetic evolution of the H9N2 avian influenza virus in Korean poultry farms. Virus Genes,45, 38-47.
Lee, C. W., D. A. Senne and D. L. Suarez, 2004: Effect of vaccine use in the evolution of Mexican lineage H5N2 avian influenza virus. Journal of Virology,78, 8372-8381.
Lee, C. W., C. S. Song, Y. J. Lee, I. P. Mo, M. Garcia, D. L. Suarez and S. J. Kim, 2000: Sequence analysis of the hemagglutinin gene of H9N2 Korean avian influenza viruses and assessment of the pathogenic potential of isolate MS96. Avian Diseases, 527-535.
Lee, D. H., J. K. Park, S. S. Yuk, T. O. Erdene-Ochir, J. H. Kwon, J. B. Lee, S. Y. Park, I. S. Choi and C. S. Song, 2013: Complete genome sequence of a natural recombinant H9N2 influenza virus from wild birds in Republic of Korea. Genome announcements,1, e00159-00112.
Lee, D. H., J. K. Park, S. S. Yuk, T. O. Erdene-Ochir, J. H. Kwon, J. B. Lee, S. Y. Park, I. S. Choi and C. S. Song, 2013: Complete genome sequence of a natural recombinant H9N2 influenza virus isolated from a white-fronted goose (Anser albifrons) in South Korea. Genome Announcements,1, e00149-00113.
Lee, H. J., J. S. Kwon, D. H. Lee, Y. N. Lee, H. N. Youn, Y. J. Lee, M. C. Kim, O. M. Jeong, H. M. Kang and J. H. Kwon, 2010: Continuing evolution and interspecies transmission of influenza viruses in live bird markets in Korea. Avian Diseases,54, 738-748.
Lee, Y. N., D. H. Lee, J. K. Park, T. H. Lim, H. N. Youn, S. S. Yuk, Y. J. Lee, I. P. Mo, H. W. Sung and J. B. Lee, 2011: Isolation and characterization of a novel H9N2 influenza virus in Korean native chicken farm. Avian Diseases,55, 724-727.
Leymarie, O., C. Embury Hyatt, C. Chevalier, L. Jouneau, M. Moroldo, B. Da Costa, Y. Berhane, B. Delmas, H. M. Weingartl and R. Le Goffic, 2014: PB1-F2 attenuates virulence of highly pathogenic avian H5N1 influenza virus in chickens. Plos One,9,
Leymarie, O., G. Jouvion, P. L. Herve, C. Chevalier, V. Lorin, J. Lecardonnel, B. Da Costa, B. Delmas, N. Escriou and R. Le Goffic, 2013: Kinetic characterization of PB1-F2-mediated immunopathology during highly pathogenic avian H5N1 influenza virus infection. PLoS One,8,
Li, C., K. Yu, G. Tian, D. Yu, L. Liu, B. Jing, J. Ping and H. Chen, 2005: Evolution of H9N2 influenza viruses from domestic poultry in Mainland China. Virology,340, 70-83.
Li, S., Z. Shi, P. Jiao, G. Zhang, Z. Zhong, W. Tian, L.-P. Long, Z. Cai, X. Zhu and M. Liao, 2010: Avian-origin H3N2 canine influenza A viruses in Southern China. Infection, Genetics and Evolution,10, 1286-1288.
Li, X., W. Qi, J. He, Z. Ning, Y. Hu, J. Tian, P. Jiao, C. Xu, J. Chen and J. Richt, 2012: Molecular basis of efficient replication and pathogenicity of H9N2 avian influenza viruses in mice. PLoS One,7,
Li, X., J. Shi, J. Guo, G. Deng, Q. Zhang, J. Wang, X. He, K. Wang, J. Chen and Y. Li, 2014: Genetics, receptor binding property, and transmissibility in mammals of naturally isolated H9N2 avian influenza viruses. PLoS Pathogens,10,
Lin, D., J. Lan and Z. Zhang, 2007: Structure and function of the NS1 protein of influenza A virus. Acta Biochimica et Biophysica Sinica,39, 155-162.
Lin, P. H., T. L. Chao, S. W. Kuo, J. T. Wang, C. C. Hung, H. C. Lin, Z. Y. Yang, S. Y. Ho, C. K. Chang and M. S. Huang, 2014: Virological, serological, and antiviral studies in an imported human case of avian influenza A (H7N9) virus in Taiwan. Clinical Infectious Diseases,58, 242-246.
Lin, Y. P., M. Shaw, V. Gregory, K. Cameron, W. Lim, A. Klimov, K. Subbarao, Y. Guan, S. Krauss and K. Shortridge, 2000: Avian-to-human transmission of H9N2 subtype influenza A viruses: relationship between H9N2 and H5N1 human isolates. Proceedings of the National Academy of Sciences,97, 9654-9658.
Lin, Z., C. Xu, B. Liu, Y. Ji, Y. Fu, J. Guo and Q. Zhu, 2014: Analysis of the phylogeny of Chinese H9N2 avian influenza viruses and their pathogenicity in mice. Archives of Virology,159, 2575-2586.
Linster, M., S. van Boheemen, M. de Graaf, E. J. A. Schrauwen, P. Lexmond, B. Mänz, T. M. Bestebroer, J. Baumann, D. van Riel and G. F. Rimmelzwaan, 2014: Identification, characterization, and natural selection of mutations driving airborne transmission of A/H5N1 virus. Cell,157, 329-339.
Liu, B., F. Havers, E. Chen, Z. Yuan, H. Yuan, J. Ou, M. Shang, K. Kang, K. Liao and F. Liu, 2014: Risk factors for influenza A (H7N9) disease—China, 2013. Clinical Infectious Diseases,59, 787-794.
Liu, H., L. Golebiewski, E. C. Dow, R. M. Krug, R. T. Javier and A. P. Rice, 2010: The ESEV PDZ-binding motif of the avian influenza A virus NS1 protein protects infected cells from apoptosis by directly targeting Scribble. Journal of Virology,84, 11164-11174.
Liu, H., X. Liu, J. Cheng, D. Peng, L. Jia and Y. Huang, 2003: Phylogenetic analysis of the hemagglutinin genes of twenty-six avian influenza viruses of subtype H9N2 isolated from chickens in China during 1996–2001. Avian Diseases,47, 116-127.
Liu, J., K. Okazaki, H. Ozaki, Y. Sakoda, Q. Wu, F. Chen and H. Kida, 2003: H9N2 influenza viruses prevalent in poultry in China are phylogenetically distinct from A/quail/Hong Kongl/G1/97 presumed to be the donor of the internal protein genes of the H5N1 Hong Kong/97 virus. Avian Pathology,32, 551-560.
Liu, J. H., K. Okazaki, W. M. Shi and H. Kida, 2003: Phylogenetic analysis of hemagglutinin and neuraminidase genes of H9N2 viruses isolated from migratory ducks. Virus Genes,27, 291-296.
Liu, J.-H., K. Okazaki, W.-M. Shi, Q.-M. Wu, A. S. Mweene and H. Kida, 2003: Phylogenetic analysis of neuraminidase gene of H9N2 influenza viruses prevalent in chickens in China during 1995–2002. Virus Genes,27, 197-202.
Liu, L., L. Zi, Z. JianFang, Z. Yun, D. Jie, Z. Xiang, G. Jun Feng and S. Yue Long, 2011: Pathogenesis and immunogenicity of an avian H9N2 influenza virus isolated from human. Biomedical and Environmental Sciences,24, 530-536.
Liu, M., Y. Guan, M. Peiris, S. He, R. J. Webby, D. Perez and R. G. Webster, 2003: The quest of influenza A viruses for new hosts. Avian Diseases,47, 849-856.
Liu, Q., H. Chen, J. Huang, Y. Chen, M. Gu, X. Wang, S. Hu, X. Liu and X. Liu, 2014: A nonpathogenic duck-origin H9N2 influenza A virus adapts to high pathogenicity in mice. Archives of Virology,159, 2243-2252.
Liu, Q., J. Huang, Y. Chen, H. Chen, Q. Li, L. He, X. Hao, J. Liu, M. Gu and J. Hu, 2015: Virulence determinants in the PB2 gene of a mouse-adapted H9N2 virus. Journal of Virology,89, 877-882.
Liu, Y., E. J. Lu, Y. L. Wang, B. Di, T. G. Li, Y. Zhou, L. L. Yang, X. Y. Xu, C. X. Fu and M. Wang, 2009: Avian influenza virus infection in people occupied in poultry fields in Guangzhou city. Zhonghua liu xing bing xue za zhi,30, 1111-1113.
Llompart, C. M., A. Nieto and A. Rodriguez-Frandsen, 2014: Specific residues of PB2 and PA influenza virus polymerase subunits confer the ability for RNA polymerase II degradation and virus pathogenicity in mice. Journal of Virology,88, 3455-3463.
Lo, Y. C., W. C. Chen, W. T. Huang, Y. C. Lin, M. C. Liu, H. W. Kuo, J. H. Chuang, J. R. Yang, M. T. Liu and H. S. Wu, 2013: Surveillance of avian influenza A (H7N9) virus infection in humans and detection of the first imported human case in Taiwan, 3 April to 10 May 2013. Euro Surveillance,18,
Lopez-Martinez, I., A. Balish, G. Barrera-Badillo, J. Jones, T. E. Nuñez-García, Y. Jang, R. Aparicio-Antonio, E. Azziz-Baumgartner, J. A. Belser and J. E. Ramirez-Gonzalez, 2013: Highly pathogenic avian influenza A (H7N3) virus in poultry workers, Mexico, 2012. Emerging Infectious Diseases,19,
Lu, J. H., X. F. Liu, W. X. Shao, Y. L. Liu, D. P. Wei and H. Q. Liu, 2005: Phylogenetic analysis of eight genes of H9N2 subtype influenza virus: a mainland China strain possessing early isolates’ genes that have been circulating. Virus Genes,31, 163-169.
Lu, S., X. Xi, Y. Zheng, Y. Cao, X. Liu and H. Lu, 2013: Analysis of the clinical characteristics and treatment of two patients with avian influenza virus (H7N9). Bioscience Trends,7, 109-112.
Lu, S., Y. Zheng, T. Li, Y. Hu, X. Liu, X. Xi, Q. Chen, Q. Wang, Y. Cao and Y. Wang, 2013: Clinical findings for early human cases of influenza A (H7N9) virus infection, Shanghai, China. Emerging Infectious Diseases,19.
Qian, M. and Y. X. Shi, 2013: H7N9 viral pneumonia: asymmetric and irregular changes on computed tomography. Chinese Medical Journal,126, 4813-4813.
Maines, T. R., X. H. Lu, S. M. Erb, L. Edwards, J. Guarner, P. W. Greer, D. C. Nguyen, K. J. Szretter, L.-M. Chen and P. Thawatsupha, 2005: Avian influenza (H5N1) viruses isolated from humans in Asia in 2004 exhibit increased virulence in mammals. Journal of Virology,79, 11788-11800.
Makarova, N. V., H. Ozaki, H. Kida, R. G. Webster and D. R. Perez, 2003: Replication and transmission of influenza viruses in Japanese quail. Virology,310, 8-15.
Malik Peiris, J. S., 2009: Avian influenza viruses in humans. Revue Scientifique et Technique,28, 161-173.
Mänz, B., M. Schwemmle and L. Brunotte, 2013: Adaptation of avian influenza A virus polymerase in mammals to overcome the host species barrier. Journal of Virology,87, 7200-7209.
Marc, D., 2014: Influenza virus non-structural protein NS1: interferon antagonism and beyond. Journal of General Virology,95, 2594-2611.
Marjuki, H., V. P. Mishin, A. P. Chesnokov, J. Jones, J. A. De La Cruz, K. Sleeman, D. Tamura, H. T. Nguyen, H. S. Wu and F. Y. Chang, 2015: Characterization of drug-resistant influenza A (H7N9) variants isolated from an oseltamivir-treated patient in Taiwan. Journal of Infectious Diseases,211, 249-257.
Marjuki, H., C. Scholtissek, J. Franks, N. J. Negovetich, J. R. Aldridge, R. Salomon, D. Finkelstein and R. G. Webster, 2010: Three amino acid changes in PB1-F2 of highly pathogenic H5N1 avian influenza virus affect pathogenicity in mallard ducks. Archives of Virology,155, 925-934.
Markwell, D. D. and K. F. Shortridge, 1982: Possible waterborne transmission and maintenance of influenza viruses in domestic ducks. Applied and Environmental Microbiology,43, 110-115.
Mase, M., T. Imada, Y. Sanada, M. Etoh, N. Sanada, K. Tsukamoto, Y. Kawaoka and S. Yamaguchi, 2001: Imported parakeets harbor H9N2 influenza A viruses that are genetically closely related to those transmitted to humans in Hong Kong. Journal of Virology,75, 3490-3494.
Matrosovich, M. N., S. Krauss and R. G. Webster, 2001: H9N2 influenza A viruses from poultry in Asia have human virus-like receptor specificity. Virology,281, 156-162.
McAuley, J. L., J. E. Chipuk, K. L. Boyd, N. Van De Velde, D. R. Green and J. A. McCullers, 2010: PB1-F2 proteins from H5N1 and 20th century pandemic influenza viruses cause immunopathology. PLoS Pathogens,6,
McAuley, J. L., F. Hornung, K. L. Boyd, A. M. Smith, R. McKeon, J. Bennink, J. W. Yewdell and J. A. McCullers, 2007: Expression of the 1918 influenza A virus PB1-F2 enhances the pathogenesis of viral and secondary bacterial pneumonia. Cell Host & Microbes,2, 240-249.
McAuley, J. L., K. Zhang and J. A. McCullers, 2010: The effects of influenza A virus PB1-F2 protein on polymerase activity are strain specific and do not impact pathogenesis. Journal of Virology,84, 558-564.
McAuley, J. 2014. International Commintte on Taxonomy of Viruses. www.ictvonline.org.
Mehle, A. and J. A. Doudna, 2009: Adaptive strategies of the influenza virus polymerase for replication in humans. Proceedings of the National Academy of Sciences,106, 21312-21316.
Mok, C. K. P., H. H. Y. Lee, M. Lestra, J. M. Nicholls, M. C. W. Chan, S. F. Sia, H. Zhu, L. L. M. Poon, Y. Guan and J. S. M. Peiris, 2014: Amino acid substitutions in polymerase basic protein 2 gene contribute to the pathogenicity of the novel A/H7N9 influenza virus in mammalian hosts. Journal of Virology,88, 3568-3576.
Mok, C. K. P., H. L. Yen, M. Y. M. Yu, K. M. Yuen, S. F. Sia, M. C. W. Chan, G. Qin, W. W. Tu and J. S. M. Peiris, 2011: Amino acid residues 253 and 591 of the PB2 protein of avian influenza virus A H9N2 contribute to mammalian pathogenesis. Journal of Virology,85, 9641-9645.
Monne, I., H. A. Hussein, A. Fusaro, V. Valastro, M. M. Hamoud, R. A. Khalefa, S. N. Dardir, M. I. Radwan, I. Capua and G. Cattoli, 2013: H9N2 influenza A virus circulates in H5N1 endemically infected poultry population in Egypt. Influenza Other Respiratory Viruses,7, 240-243.
Mullaney, R., 2003: Live-bird market closure activities in the northeastern United States. Avian Diseases,47, 1096-1098.
Munir, M., S. Zohari, M. Abbas, M. Z. Shabbir, M. N. Zahid, M. S. Latif, M. Iqbal, Z. U. A. Qureshi and M. Berg, 2013: Isolation and characterization of low pathogenic H9N2 avian influenza A viruses from a healthy flock and its comparison to other H9N2 isolates. Indian Journal of Virology,24, 342-348.
Muramoto, Y., T. Noda, E. Kawakami, R. Akkina and Y. Kawaoka, 2013: Identification of novel influenza A virus proteins translated from PA mRNA. Journal of Virology,87, 2455-2462.
Naeem, K., A. Ullah, R. Manvell and D. J. Alexander, 1999: Avian influenza A subtype H9N2 in poultry in Pakistan. Veterinary record,145,
Nagarajan, S., K. Rajukumar, C. Tosh, V. Ramaswamy, K. Purohit, G. Saxena, P. Behera, B. Pattnaik, H. K. Pradhan and S. C. Dubey, 2009: Isolation and pathotyping of H9N2 avian influenza viruses in Indian poultry. Veterinary Microbiology,133, 154-163.
Nailwal, H., K. Kamra and S. K. Lal, 2014: H7N9: A killer in the making or a false alarm? Canadian Journal of Microbiology,60, 425-429.
Narayan, O., G. Lang and B. Rouse, 1969: A new influenza A virus infection in turkeys. IV. Experimental susceptibility of domestic birds to virus strain turkey/Ontario 7732/1966. Archiv für die gesamte Virusforschung,26, 149-165.
Negovetich, N. J., M. M. Feeroz, L. Jones-Engel, D. Walker, S. R. Alam, K. Hasan, P. Seiler, A. Ferguson, K. Friedman and S. Barman, 2011: Live bird markets of Bangladesh: H9N2 viruses and the near absence of highly pathogenic H5N1 influenza. Plos One,6,
Neumann, G., M. T. Hughes and Y. Kawaoka, 2000: Influenza A virus NS2 protein mediates vRNP nuclear export through NES‐independent interaction with hCRM1. The EMBO Journal,19, 6751-6758.
Nicholson, K. G., C. I. Thompson, J. M. Klap, J. M. Wood, S. Batham, R. W. Newman, R. Mischler, M. C. Zambon and I. Stephenson, 2009: Safety and immunogenicity of whole-virus, alum-adjuvanted whole-virus, virosomal, and whole-virus intradermal influenza A/H9N2 vaccine formulations. Vaccine,28, 171-178.
Nili, H. and K. Asasi, 2003: Avian influenza (H9N2) outbreak in Iran. Avian Diseases,47, 828-831.
Nili, H. and K. Asasi, 2002: Natural cases and an experimental study of H9N2 avian influenza in commercial broiler chickens of Iran. Avian Pathology,31, 247-252.
Nili, H., K. Asasi, H. Dadras and M. Ebrahimi, 2007: Pathobiology of H9N2 avian influenza virus in Japanese quail (Coturnix coturnix japonica). Avian Diseases,51, 390-392.
Nili, H., A. Mohammadi, H. Habibi and S. Firouzi, 2013: Pathogenesis of H9N2 virus in Chukar partridges. Avian Pathology,42, 230-234.
O’Neill, R. E., J. Talon and P. Palese, 1998: The influenza virus NEP (NS2 protein) mediates the nuclear export of viral ribonucleoproteins. The EMBO Journal,17, 288-296.
Odagiri, T., K. Tominaga, K. Tobita and S. Ohta, 1994: An amino acid change in the non-structural NS2 protein of an influenza A virus mutant is responsible for the generation of defective interfering (DI) particles by amplifying DI RNAs and suppressing complementary RNA synthesis. Journal of General Virology,75, 43-53.
Ohtsu, Y., Y. Honda, Y. Sakata, H. Kato and T. Toyoda, 2002: Fine mapping of the subunit binding sites of influenza virus RNA polymerase. Microbiology and Immunology,46, 167-175.
Okamatsu, M., T. Saito, M. Mase, K. Tsukamoto and S. Yamaguchi, 2007: Characterization of H5N2 influenza A viruses isolated from chickens in Japan. Avian Diseases,51, 474-475.
Okamatsu, M., T. Saito, Y. Yamamoto, M. Mase, S. Tsuduku, K. Nakamura, K. Tsukamoto and S. Yamaguchi, 2007: Low pathogenicity H5N2 avian influenza outbreak in Japan during the 2005–2006. Veterinary Microbiology,124, 35-46.
Pantin-Jackwood, M. J., P. J. Miller, E. Spackman, D. E. Swayne, L. Susta, M. Costa-Hurtado and D. L. Suarez, 2014: Role of poultry in the spread of novel H7N9 influenza virus in China. Journal of Virology,88, 5381-5390.
Parvin, R., K. Heenemann, M. Y. Halami, E. H. Chowdhury, M. Islam and T. W. Vahlenkamp, 2014: Full-genome analysis of avian influenza virus H9N2 from Bangladesh reveals internal gene reassortments with two distinct highly pathogenic avian influenza viruses. Archives of Virology,159, 1651-1661.
Pawar, S. D., B. V. Tandale, C. G. Raut, S. S. Parkhi, T. D. Barde, Y. K. Gurav, S. S. Kode and A. C. Mishra, 2012: Avian influenza H9N2 seroprevalence among poultry workers in Pune, India, 2010. Plos One,7,
Peiris, M., K. Y. Yuen, C. W. Leung, K. H. Chan, P. L. S. Ip, R. W. M. Lai, W. K. Orr and K. F. Shortridge, 1999: Human infection with influenza H9N2. The Lancet,354, 916-917.
Pena, L., A. L. Vincent, C. L. Loving, J. N. Henningson, K. M. Lager, W. Li and D. R. Perez, 2012: Strain-dependent effects of PB1-F2 of triple-reassortant H3N2 influenza viruses in swine. Journal of General Virology,
Pena, L., A. L. Vincent, C. L. Loving, J. N. Henningson, K. M. Lager, A. Lorusso and D. R. Perez, 2012: Restored PB1-F2 in the 2009 pandemic H1N1 influenza virus has minimal effects in swine. Journal of Virology,86, 5523-5532.
Pérez, D. R. and R. O. Donis, 1995: A 48-amino-acid region of influenza A virus PB1 protein is sufficient for complex formation with PA. Journal of Virology,69, 6932-6939.
Perez, D. R. and R. O. Donis, 2001: Functional analysis of PA binding by influenza a virus PB1: effects on polymerase activity and viral infectivity. Journal of Virology,75, 8127-8136.
Perez, D. R., W. Lim, J. P. Seiler, G. Yi, M. Peiris, K. F. Shortridge and R. G. Webster, 2003: Role of quail in the interspecies transmission of H9 influenza A viruses: molecular changes on HA that correspond to adaptation from ducks to chickens. Journal of Virology,77, 3148-3156.
Perez, D. R., R. J. Webby, E. Hoffmann and R. G. Webster, 2003: Land-based birds as potential disseminators of avian/mammalian reassortant influenza A viruses. Avian Diseases,47, 1114-1117.
Perk, S., C. Banet-Noach, E. Shihmanter, S. Pokamunski, M. Pirak, M. Lipkind and A. Panshin, 2006: Genetic characterization of the H9N2 influenza viruses circulated in the poultry population in Israel. Comparative Immunology, Microbiology and Infectious Diseases,29, 207-223.
Perk, S., A. Panshin, E. Shihmanter, I. Gissin, S. Pokamunski, M. Pirak and M. Lipkind, 2006: Ecology and molecular epidemiology of H9N2 avian influenza viruses isolated in Israel during 2000-2004 epizootic. Developments in Biologicals,124, 201-209.
Poole, E., D. Elton, L. Medcalf and P. Digard, 2004: Functional domains of the influenza A virus PB2 protein: identification of NP-and PB1-binding sites. Virology,321, 120-133.
Qi, W., X. Zhou, W. Shi, L. Huang, W. Xia, D. Liu, H. Li, S. Chen, F. Lei and L. Cao, 2014: Genesis of the novel human-infecting influenza A (H10N8) virus and potential genetic diversity of the virus in poultry, China. Euro Surveillance,19,
Qiao, C., Q. Liu, B. Bawa, H. Shen, W. Qi, Y. Chen, C. K. P. Mok, A. García-Sastre, J. A. Richt and W. Ma, 2012: Pathogenicity and transmissibility of reassortant H9 influenza viruses with genes from pandemic H1N1 virus. Journal of General Virology,93,
Reuther, P., B. Mänz, L. Brunotte, M. Schwemmle and K. Wunderlich, 2011: Targeting of the influenza A virus polymerase PB1-PB2 interface indicates strain-specific assembly differences. Journal of Virology,85, 13298-13309.
Richard, M., E. J. Schrauwen, M. de Graaf, T. M. Bestebroer, M. I. Spronken, S. van Boheemen, D. de Meulder, P. Lexmond, M. Linster and S. Herfst, 2013: Limited airborne transmission of H7N9 influenza A virus between ferrets. Nature,501, 560-563.
Riegger, D., R. Hai, D. Dornfeld, B. Mänz, V. Leyva-Grado, M. T. Sánchez-Aparicio, R. A. Albrecht, P. Palese, O. Haller and M. Schwemmle, 2015: The nucleoprotein of newly emerged H7N9 influenza A virus harbors a unique motif conferring resistance to antiviral human MxA. Journal of Virology,89, 2241-2252.
Rolling, T., I. Koerner, P. Zimmermann, K. Holz, O. Haller, P. Staeheli and G. Kochs, 2009: Adaptive mutations resulting in enhanced polymerase activity contribute to high virulence of influenza A virus in mice. Journal of Virology,83, 6673-6680.
Rui-Hua, Z., C. Hong-Yu, X. Ming-Ju, L. Kai, C. Hua-Lan, W. Cun-Lian, W. Dong, L. Cun-Xin and X. Tong, 2011: Molecular characterization and pathogenicity of swine influenza H9N2 subtype virus A/swine/HeBei/012/2008/(H9N2). Acta Neurologica 55, 219-226.
Saito, T., W. Lim, T. Suzuki, Y. Suzuki, H. Kida, S. Nishimura and M. Tashiro, 2001: Characterization of a human H9N2 influenza virus isolated in Hong Kong. Vaccine,20, 125-133.
Sakai, K., Y. Ami, M. Tahara, T. Kubota, M. Anraku, M. Abe, N. Nakajima, T. Sekizuka, K. Shirato and Y. Suzaki, 2014: The host protease TMPRSS2 plays a major role in in vivo replication of emerging H7N9 and seasonal influenza viruses. Journal of Virology,88, 5608-5616.
Saw, T. A., W. Lim, K. F. Shortridge, J. Tam, K. K. Liu, K. H. Mak, T. Tsang, F. Y. Lee and H. Kwong, 1997: Isolation of avian influenza A (H5N1) viruses from humans–Hong Kong, May-December 1997. Morbidity and Mortality Weekly Report,46, 1204-1207.
Schmolke, M., B. Manicassamy, L. Pena, T. Sutton, R. Hai, Z. T. Varga, B. G. Hale, J. Steel, D. R. Perez and A. García-Sastre, 2011: Differential contribution of PB1-F2 to the virulence of highly pathogenic H5N1 influenza A virus in mammalian and avian species. PLoS Pathogens,7,
Scholtissek, C. and E. Naylor, 1988: Fish farming and influenza pandemics. Nature,331, 215-215.
Seyer, R., E. R. Hrincius, D. Ritzel, M. Abt, A. Mellmann, H. Marjuki, J. Kühn, T. Wolff, S. Ludwig and C. Ehrhardt, 2012: Synergistic adaptive mutations in the hemagglutinin and polymerase acidic protein lead to increased virulence of pandemic 2009 H1N1 influenza A virus in mice. Journal of Infectious Diseases,205, 262-271.
Shahsavandi, S., A.-H. Salmanian, S. A. Ghorashi, S. Masoudi and M. M. Ebrahimi, 2012: Evolutionary characterization of hemagglutinin gene of H9N2 influenza viruses isolated from Asia. Research in Veterinary Science,93, 234-239.
Shepard, S. S., C. T. Davis, J. Bahl, P. Rivailler, I. A. York and R. O. Donis, 2014: LABEL: fast and accurate lineage assignment with assessment of H5N1 and H9N2 influenza A hemagglutinins. Plos One,9,
Shi, M., B. W. Jagger, H. M. Wise, P. Digard, E. C. Holmes and J. K. Taubenberger, 2012: Evolutionary conservation of the PA-X open reading frame in segment 3 of influenza A virus. Journal of Virology,86, 12411-12413.
Shi, Y., W. Zhang, F. Wang, J. Qi, Y. Wu, H. Song, F. Gao, Y. Bi, Y. Zhang and Z. Fan, 2013: Structures and receptor binding of hemagglutinins from human-infecting H7N9 influenza viruses. Science,342, 243-247.
Shortridge, K., 1992: Pandemic influenza: a zoonosis? Seminars in Respiratory Infections,7, 11-25.
Sieverding, E., 2014: Zur aktuellen Situation der H9-Infektionen bei Puten. 87th Fachgesprach Uber Geflugelkrankheiten, 51-53.
Sims, L. D., J. Domenech, C. Benigno, S. Kahn, A. Kamata, J. Lubroth, V. Martin and P. Roeder, 2005: Origin and evolution of highly pathogenic H5N1 avian influenza in Asia. Veterinary Record,157, 159-164.
Slemons, R. D. and B. C. Easterday, 1972: Host response differences among 5 avian species to an influenzavirus—A/turkey/Ontario/7732/66 (Hav5N?). Bulletin of the World Health Organization,47,
Slemons, R. D. and B. C. Easterday, 1977: Type-A influenza viruses in the feces of migratory waterfowl. Journal of the American Veterinary Medical Association,171, 947-948.
Slemons, R. D. and B. C. Easterday, 1978: Virus replication in the digestive tract of ducks exposed by aerosol to type-A influenza. Avian Diseases,22, 367-377.
Smeenk, C. A. and E. G. Brown, 1994: The influenza virus variant A/FM/1/47-MA possesses single amino acid replacements in the hemagglutinin, controlling virulence, and in the matrix protein, controlling virulence as well as growth. Journal of General Virology,68, 530-534.
Smietanka, K. and Z. Minta, 2014: Avian influenza in Poland. Acta Biochimica Polonica,61, 453-457.
Śmietanka, K., Z. Minta, E. Świętoń, M. Olszewska, M. Jóźwiak, K. Domańska-Blicharz, K. Wyrostek, G. Tomczyk and A. Pikuła, 2014: Avian influenza H9N2 subtype in Poland–characterization of the isolates and evidence of concomitant infections. Avian Pathology,43, 427-436.
Smithies, L. K., D. B. Radloff, R. W. Friedell, G. W. Albright, V. E. Misner and B. C. Easterday, 1969: Two different type A influenza virus infections in turkeys in Wisconsin I. 1965-66 outbreak. Avian Diseases,13, 603-606.
Soltanialvar, M., H. Shoushtari, M. Bozorgmehrifard, S. Charkhkar and F. Akbarnejad, 2012: Sequence and phylogenetic analysis of neuraminidase genes of H9N2 avian influenza viruses isolated from commercial broiler chicken in Iran (2008 and 2009). Tropical Animal Health and Production,44, 419-425.
Song, D., H.-J. Moon, D.-J. An, H.-Y. Jeoung, H. Kim, M.-J. Yeom, M. Hong, J.-H. Nam, S.-J. Park and B.-K. Park, 2012: A novel reassortant canine H3N1 influenza virus between pandemic H1N1 and canine H3N2 influenza viruses in Korea. Journal of General Virology,93,
Song, H., H. Wan, Y. Araya and D. R. Perez, 2009: Partial direct contact transmission in ferrets of a mallard H7N3 influenza virus with typical avian-like receptor specificity. Virology Journal,6, 1-12.
Song, P., J. Xia, J. Gao, L. Xu, Y. Huang, L. Yao and W. Tang, 2013: Measures to combat H7N9 virus infection in China: live poultry purchasing habits, poultry handling, and living conditions increase the risk of exposure to contaminated environments. Bioscience Trends,7, 168-171.
Sorrell, E. M. and D. R. Perez, 2007: Adaptation of influenza A/Mallard/Potsdam/178-4/83 H2N2 virus in Japanese quail leads to infection and transmission in chickens. Avian Diseases,51, 264-268.
Sorrell, E. M., H. Wan, Y. Araya, H. Song and D. R. Perez, 2009: Minimal molecular constraints for respiratory droplet transmission of an avian–human H9N2 influenza A virus. Proceedings of the National Academy of Sciences,106, 7565-7570.
Soubies, S. M., T. W. Hoffmann, G. Croville, T. Larcher, M. Ledevin, D. Soubieux, P. Quéré, J.-L. Guérin, D. Marc and R. Volmer, 2013: Deletion of the C-terminal ESEV domain of NS1 does not affect the replication of a low-pathogenic avian influenza virus H7N1 in ducks and chickens. Journal of General Virology,94, 50-58.
Spesock, A., M. Malur, M. J. Hossain, L.-M. Chen, B. L. Njaa, C. T. Davis, A. S. Lipatov, I. A. York, R. M. Krug and R. O. Donis, 2011: The virulence of 1997 H5N1 influenza viruses in the mouse model is increased by correcting a defect in their NS1 proteins. Journal of Virology,85, 7048-7058.
Stephenson, I., K. G. Nicholson, R. Glück, R. Mischler, R. W. Newman, A. M. Palache, N. Q. Verlander, F. Warburton, J. M. Wood and M. C. Zambon, 2003: Safety and antigenicity of whole virus and subunit influenza A/Hong Kong/1073/99 (H9N2) vaccine in healthy adults: phase I randomised trial. The Lancet,362, 1959-1966.
Suarez, D. L., M. Garcia, J. Latimer, D. Senne and M. Perdue, 1999: Phylogenetic analysis of H7 avian influenza viruses isolated from the live bird markets of the Northeast United States. Journal of Virology,73, 3567-3573.
Subbarao, E. K., W. London and B. R. Murphy, 1993: A single amino acid in the PB2 gene of influenza A virus is a determinant of host range. Journal of Virology,67, 1761-1764.
Sun, X., X. Xu, Q. Liu, D. Liang, C. Li, Q. He, J. Jiang, Y. Cui, J. Li and L. Zheng, 2013: Evidence of avian-like H9N2 influenza A virus among dogs in Guangxi, China. Infection, Genetics and Evolution,20, 471-475.
Sun, Y. and J. Liu, 2015: H9N2 influenza virus in China: a cause of concern. Protein and Cell,6, 18-25.
Sun, Y., J. Pu, Z. Jiang, T. Guan, Y. Xia, Q. Xu, L. Liu, B. Ma, F. Tian and E. G. Brown, 2010: Genotypic evolution and antigenic drift of H9N2 influenza viruses in China from 1994 to 2008. Veterinary Microbiology,146, 215-225.
Sun, Y., K. Qin, J. Wang, J. Pu, Q. Tang, Y. Hu, Y. Bi, X. Zhao, H. Yang and Y. Shu, 2011: High genetic compatibility and increased pathogenicity of reassortants derived from avian H9N2 and pandemic H1N1/2009 influenza viruses. Proceedings of the National Academy of Sciences,108, 4164-4169.
Sun, Y., Y. Tan, K. Wei, H. Sun, Y. Shi, J. Pu, H. Yang, G. F. Gao, Y. Yin and W. Feng, 2013: Amino acid 316 of hemagglutinin and the neuraminidase stalk length influence virulence of H9N2 influenza virus in chickens and mice. Journal of Virology,87, 2963-2968.
Sun, Y., Q. Xu, Y. Shen, L. Liu, K. Wei, H. Sun, J. Pu, K.-C. Chang and J. Liu, 2014: Naturally occurring mutations in the PA gene are key contributors to increased virulence of pandemic H1N1/09 influenza virus in mice. Journal of Virology,88, 4600-4604.
Sutton, T. C., C. Finch, H. Shao, M. Angel, H. Chen, I. Capua, G. Cattoli, I. Monne and D. R. Perez, 2014: Airborne transmission of highly pathogenic H7N1 influenza virus in ferrets. Journal of Virology,88, 6623-6635.
Tada, T., K. Suzuki, Y. Sakurai, M. Kubo, H. Okada, T. Itoh and K. Tsukamoto, 2011: Emergence of avian influenza viruses with enhanced transcription activity by a single amino acid substitution in the nucleoprotein during replication in chicken brains. Journal of Virology,85, 10354-10363.
Tarnow, C., G. Engels, A. Arendt, F. Schwalm, H. Sediri, A. Preuss, P. S. Nelson, W. Garten, H.-D. Klenk and G. Gabriel, 2014: TMPRSS2 is a host factor that is essential for pneumotropism and pathogenicity of H7N9 influenza A virus in mice. Journal of Virology,88, 4744-4751.
Thakar, J., S. Schmid, J. L. Duke, A. García-Sastre and S. H. Kleinstein, 2013: Overcoming NS1-mediated immune antagonism involves both interferon-dependent and independent mechanisms. Journal of Interferon and Cytokine Research,33, 700-708.
Tharakaraman, K., A. Jayaraman, R. Raman, K. Viswanathan, N. W. Stebbins, D. Johnson, Z. Shriver, V. Sasisekharan and R. Sasisekharan, 2013: Glycan receptor binding of the influenza A virus H7N9 hemagglutinin. Cell,153, 1486-1493.
Tian, J., W. Qi, X. Li, J. He, P. Jiao, C. Zhang, G.-Q. Liu and M. Liao, 2012: A single E627K mutation in the PB2 protein of H9N2 avian influenza virus increases virulence by inducing higher glucocorticoids (GCs) level. Plos One,7,
To, K. K.-W., W. Song, S.-Y. Lau, T.-L. Que, D. C. Lung, I. F.-N. Hung, H. Chen and K.-Y. Yuen, 2014: Unique reassortant of influenza A (H7N9) virus associated with severe disease emerging in Hong Kong. Journal of Infection,69, 60-68.
To, K. K. W., A. K. L. Tsang, J. F. W. Chan, V. C. C. Cheng, H. Chen and K.-Y. Yuen, 2014: Emergence in China of human disease due to avian influenza A (H10N8)–cause for concern? Journal of Infection,68, 205-215.
Tosh, C., S. Nagarajan, P. Behera, K. Rajukumar, K. Purohit, R. P. Kamal, H. V. Murugkar, S. Gounalan, B. Pattnaik and P. R. Vanamayya, 2008: Genetic analysis of H9N2 avian influenza viruses isolated from India. Archives of Virology,153, 1433-1439.
Trock, S. C., D. A. Senne, M. Gaeta, A. Gonzalez and B. Lucio, 2003: Low-Pathogenicity Avian Influenza Virus in Live Bird Markets—What About the Livestock Area? Avian Diseases,47, 1111-1113.
Tse, L. V., A. M. Hamilton, T. Friling and G. R. Whittaker, 2014: A novel activation mechanism of avian influenza virus H9N2 by furin. Journal of Virology,88, 1673-1683.
Tumpey, T. M., D. R. Kapczynski and D. E. Swayne, 2004: Comparative susceptibility of chickens and turkeys to avian influenza A H7N2 virus infection and protective efficacy of a commercial avian influenza H7N2 virus vaccine. Avian Diseases,48, 167-176.
Tweed, S. A., D. M. Skowronski, S. T. David, A. Larder, M. Petric, W. Lees, Y. Li, J. Katz, M. Krajden and R. Tellier, 2004: Human illness from avian influenza H7N3, British Columbia. Emerging Infectious Diseases,10, 2196-2199.
Uyeki, T. M., Y.-H. Chong, J. M. Katz, W. Lim, Y.-Y. Ho, S. S. Wang, T. H. Tsang, W. W.-Y. Au, S.-C. Chan and T. Rowe, 2002: Lack of evidence for human-to-human transmission of avian influenza A (H9N2) viruses in Hong Kong, China 1999. Emerging Infectious Diseases,8,
Uyeki, T. M., D. C. Nguyen, T. Rowe, X. Lu, J. Hu-Primmer, L. P. Huynh, N. L. Hang and J. M. Katz, 2012: Seroprevalence of antibodies to avian influenza A (H5) and A (H9) viruses among market poultry workers, Hanoi, Vietnam, 2001. Plos One,7,
Van Den Berg, T. and P. Houdart, 2008: Avian influenza outbreak management: action at time of confirmation, depopulation and disposal methods; the ‘Belgian experience’during the H7N7 highly pathogenic avian influenza epidemic in 2003. Zoonoses and Public Health,55, 54-64.
Vasin, A. V., O. A. Temkina, V. V. Egorov, S. A. Klotchenko, M. A. Plotnikova and O. I. Kiselev, 2014: Molecular mechanisms enhancing the proteome of influenza A viruses: an overview of recently discovered proteins. Virus Research,185, 53-63.
Villarreal, C., 2009: Avian influenza in Mexico. Revue Scientifique et Technique,28, 261-265.
Wan, H. and D. R. Perez, 2007: Amino acid 226 in the hemagglutinin of H9N2 influenza viruses determines cell tropism and replication in human airway epithelial cells. Journal of Virology,81, 5181-5191.
Wan, H. and D. R. Perez, 2006: Quail carry sialic acid receptors compatible with binding of avian and human influenza viruses. Journal of Virology,346, 278-286.
Wan, H., E. M. Sorrell, H. Song, M. J. Hossain, G. Ramirez-Nieto, I. Monne, J. Stevens, G. Cattoli, I. Capua and L.-M. Chen, 2008: Replication and transmission of H9N2 influenza viruses in ferrets: evaluation of pandemic potential. Plos One,3,
Wang, B., Q. Chen and Z. Chen, 2012: Complete genome sequence of an H9N2 avian influenza virus isolated from egret in Lake Dongting wetland. Journal of Virology,86,
Wang, J., Y. Sun, Q. Xu, Y. Tan, J. Pu, H. Yang, E. G. Brown and J. Liu, 2012: Mouse-adapted H9N2 influenza A virus PB2 protein M147L and E627K mutations are critical for high virulence. Plos One,7,
Wang, S. Y., S. H. Ren, M. X. Huang, D. J. Yu, S. Qiang, H. F. Zhao, Q. H. Lü and S. X. Qian, 2013: Clinical variability in onset of influenza A (H7N9) infection. Chinese Medical Journal,126, 4194-4196.
Wang, X. F., G. C. Shi, H. Y. Wan, S. G. Hang, H. Chen, W. Chen, H. P. Qu, B. H. Han and M. Zhou, 2014: Clinical features of three avian influenza H7N 9 virus‐infected patients in Shanghai. The Clinical Respiratory Journal,8, 410-416.
Wang, Y., Y. Zhang, H. Wang, Z. Zhou, Z. Zhou, Q. Gu, C. Wu and B. Zhu, 2013: Sharing insights and H7N9 patient clinical data. American Journal of Respiratory and Critical Care Medicine,188, 115-117.
Wasilenko, J. L., L. Sarmento and M. J. Pantin-Jackwood, 2009: A single substitution in amino acid 184 of the NP protein alters the replication and pathogenicity of H5N1 avian influenza viruses in chickens. Archives of Virology,154, 969-979.
Webster, R. G., W. J. Bean, O. T. Gorman, T. M. Chambers and Y. Kawaoka, 1992: Evolution and ecology of influenza A viruses. Microbiological Reviews,56, 152-179.
Webster, R. G., V. S. Hinshaw, W. J. Bean Jr, B. Turner and K. F. Shortridge, 1977: Influenza viruses from avian and porcine sources and their possible role in the origin of human pandemic strains. Developments in Biological Standardization,39, 461-468.
Webster, R. G., M. Yakhno, V. S. Hinshaw, W. J. Bean and K. C. Murti, 1978: Intestinal influenza: replication and characterization of influenza viruses in ducks. Virology Journal,84, 268-278.
World Health Organization, 2014: Human cases of influenza at the human-animal interface, 2013. Weekly Epidemiological Record,89, 309-320.
Wise, H. M., E. C. Hutchinson, B. W. Jagger, A. D. Stuart, Z. H. Kang, N. Robb, L. M. Schwartzman, J. C. Kash, E. Fodor and A. E. Firth, 2012: Identification of a novel splice variant form of the influenza A virus M2 ion channel with an antigenically distinct ectodomain. PLoS Pathogens,8,
Wiwanitkit, V., 2013: Neurological manifestation in new emerging H7N9 influenza: an issue in neurology. Acta Neurologica Taiwanica,22, 138-139.
Wright, S. M., Y. Kawaoka, G. B. Sharp, D. A. Senne and R. G. Webster, 1992: Interspecies transmission and reassortment of influenza A viruses in pigs and turkeys in the United States. American Journal of Epidemiology,136, 488-497.
Wu, P., H. Jiang, J. T. Wu, E. Chen, J. He, H. Zhou, L. Wei, J. Yang, B. Yang and Y. Qin, 2014: Poultry market closures and human infection with influenza A (H7N9) virus, China, 2013–14. Emerging Infectious Diseases,20,
Wu, R., Z. Sui, Z. Liu, W. Liang, K. Yang, Z. Xiong and D. Xu, 2010: Transmission of avian H9N2 influenza viruses in a murine model. Veterinary Microbiology,142, 211-216.
Wu, R., H. Zhang, K. Yang, W. Liang, Z. Xiong, Z. Liu, X. Yang, H. Shao, X. Zheng and M. Chen, 2009: Multiple amino acid substitutions are involved in the adaptation of H9N2 avian influenza virus to mice. Veterinary Microbiology,138, 85-91.
Xiong, X., S. R. Martin, L. F. Haire, S. A. Wharton, R. S. Daniels, M. S. Bennett, J. W. McCauley, P. J. Collins, P. A. Walker and J. J. Skehel, 2013: Receptor binding by an H7N9 influenza virus from humans. Nature,499, 496-499.
Xu, C., W. Fan, R. Wei and H. Zhao, 2004: Isolation and identification of swine influenza recombinant A/Swine/Shandong/1/2003 (H9N2) virus. Microbes and Infection,6, 919-925.
Xu, C., W.-B. Hu, K. Xu, Y.-X. He, T.-Y. Wang, Z. Chen, T.-X. Li, J.-H. Liu, P. Buchy and B. Sun, 2012: Amino acids 473V and 598P of PB1 from an avian-origin influenza A virus contribute to polymerase activity, especially in mammalian cells. Journal of General Virology,93, 531-540.
Xu, K., L. Ferreri, A. Rimondi, V. Olivera, M. Romano, H. Ferreyra, V. Rago, M. Uhart, H. Chen and T. Sutton, 2012: Isolation and characterization of an H9N2 influenza virus isolated in Argentina. Virus Research,168, 41-47.
Xu, K., K. Li, G. Smith, J. Li, H. Tai, J. Zhang, R. Webster, J. Peiris, H. Chen and Y. Guan, 2007: Evolution and molecular epidemiology of H9N2 influenza A viruses from quail in southern China, 2000 to 2005. Journal of Virology,81, 2635-2645.
Xu, K. M., G. J. D. Smith, J. Bahl, L. Duan, H. Tai, D. Vijaykrishna, J. Wang, J. X. Zhang, K. S. Li and X. H. Fan, 2007: The genesis and evolution of H9N2 influenza viruses in poultry from southern China, 2000 to 2005. Journal of Virology,81, 10389-10401.
Xu, R., R. P. de Vries, X. Zhu, C. M. Nycholat, R. McBride, W. Yu, J. C. Paulson and I. A. Wilson, 2013: Preferential recognition of avian-like receptors in human influenza A H7N9 viruses. Science,342, 1230-1235.
Xu, X.-J., G.-Y. Xu, H.-B. Zhou, Z.-J. Yu, A.-D. Zhang, Y.-F. Song, M.-L. Jin and H.-C. Chen, 2008: Evolutionary characterization of influenza virus A/duck/Hubei/W1/2004 (H9N2) isolated from central China. Virus Genes,36, 79-83.
Yamada, S., M. Hatta, B. L. Staker, S. Watanabe, M. Imai, K. Shinya, Y. Sakai-Tagawa, M. Ito, M. Ozawa and T. Watanabe, 2010: Biological and structural characterization of a host-adapting amino acid in influenza virus. PLoS pathogens,6,
Yamayoshi, S., S. Yamada, S. Fukuyama, S. Murakami, D. Zhao, R. Uraki, T. Watanabe, Y. Tomita, C. Macken and G. Neumann, 2014: Virulence-affecting amino acid changes in the PA protein of H7N9 influenza A viruses. Journal of Virology,88, 3127-3134.
Yang, Y., F. Guo, W. Zhao, Q. Gu, M. Huang, Q. Cao, Y. Shi, J. Li, J. Chen and J. Yan, 2015: Novel avian-origin influenza A (H7N9) in critically ill patients in China. Critical Care Medicine,43, 339-345.
Ye, J., E. M. Sorrell, Y. Cai, H. Shao, K. Xu, L. Pena, D. Hickman, H. Song, M. Angel and R. A. Medina, 2010: Variations in the hemagglutinin of the 2009 H1N1 pandemic virus: potential for strains with altered virulence phenotype? PLoS Pathogens,6,
Yee, K. S., C. A. Novick, D. A. Halvorson, N. Dao, T. E. Carpenter and C. J. Cardona, 2011: Prevalence of low pathogenicity avian influenza virus during 2005 in two US live bird market systems. Avian Diseases,55, 236-242.
Yu, H., B. J. Cowling, L. Feng, E. H. Lau, Q. Liao, T. K. Tsang, Z. Peng, P. Wu, F. Liu and V. J. Fang, 2013: Human infection with avian influenza A H7N9 virus: an assessment of clinical severity. The Lancet,382, 138-145.
Yu, H., R.-H. Hua, T.-C. Wei, Y.-J. Zhou, Z.-J. Tian, G.-X. Li, T.-Q. Liu and G.-Z. Tong, 2008: Isolation and genetic characterization of avian origin H9N2 influenza viruses from pigs in China. Veterinary Microbiology,131, 82-92.
Yu, H., J. T. Wu, B. J. Cowling, Q. Liao, V. J. Fang, S. Zhou, P. Wu, H. Zhou, E. H. Lau and D. Guo, 2014: Effect of closure of live poultry markets on poultry-to-person transmission of avian influenza A H7N9 virus: an ecological study. The Lancet,383, 541-548.
Yu, H., Y.-J. Zhou, G.-X. Li, J.-H. Ma, L.-P. Yan, B. Wang, F.-R. Yang, M. Huang and G.-Z. Tong, 2011: Genetic diversity of H9N2 influenza viruses from pigs in China: a potential threat to human health? Veterinary Microbiology,149, 254-261.
Yu, J., X. Li, Y. Wang, B. Li, H. Li, Y. Li, W. Zhou, C. Zhang, Y. Wang and Z. Rao, 2011: PDlim2 selectively interacts with the PDZ binding motif of highly pathogenic avian H5N1 influenza A virus NS1. Plos One,6,
Yu, L., Z. Wang, Y. Chen, W. Ding, H. Jia, J. F.-W. Chan, K. K.-W. To, H. Chen, Y. Yang and W. Liang, 2013: Clinical, virological, and histopathological manifestations of fatal human infections by avian influenza A (H7N9) virus. Clinical Infectious Diseases,57, 1449-1457.
Yu, Z., K. Cheng, W. Sun, Y. Xin, J. Cai, R. Ma, Q. Zhao, L. Li, J. Huang and X. Sang, 2014: Lowly pathogenic avian influenza (H9N2) infection in Plateau pika (Ochotona curzoniae), Qinghai Lake, China. Veterinary Microbiology,173, 132-135.
Yu, X., T. Jin, Y. Cui, X. Pu, J. Li, J. Xu, G. Liu, H. Jia, D. Liu and S. Song, 2014: Influenza H7N9 and H9N2 viruses: coexistence in poultry linked to human H7N9 infection and genome characteristics. Journal of Virology,88, 3423-3431.
Yu, Z., K. Cheng, W. Sun, Y. Xin, J. Cai, R. Ma, Q. Zhao, L. Li, J. Huang and X. Sang, 2014: Lowly pathogenic avian influenza (H9N2) infection in Plateau pika (Ochotona curzoniae), Qinghai Lake, China. Veterinary Microbiology,173, 132-135.
Zhang, J. and Y. Chen, 2014: Patients infected with avian influenza A H7N9 virus have abnormally low thyroid hormone levels. Clinical Laboratory,60, 1241-1243.
Zhang, K., W. Xu, Z. Zhang, T. Wang, X. Sang, K. Cheng, Z. Yu, X. Zheng, H. Wang and Y. Zhao, 2013: Experimental infection of non-human primates with avian influenza virus (H9N2). Archives of Virology,158, 2127-2134.
Zhang, K., Z. Zhang, Z. Yu, L. Li, K. Cheng, T. Wang, G. Huang, S. Yang, Y. Zhao and N. Feng, 2013: Domestic cats and dogs are susceptible to H9N2 avian influenza virus. Virus Research,175, 52-57.
Zhang, P., Y. Tang, X. Liu, D. Peng, W. Liu, H. Liu, S. Lu and X. Liu, 2008: Characterization of H9N2 influenza viruses isolated from vaccinated flocks in an integrated broiler chicken operation in eastern China during a 5 year period (1998–2002). Journal of General Virology,89, 3102-3112.
Zhang, T., Y. Bi, H. Tian, X. Li, D. Liu, Y. Wu, T. Jin, Y. Wang, Q. Chen and Z. Chen, 2014: Human infection with influenza virus A (H10N8) from live poultry markets, China, 2014. Emerging Infectious Diseases,20,
Zhang, Z., W. Jianguo, Q. Kejian, L. Xiaoqing, X. Zuke, S. Jian, Z. Zhenguo, W. Qi, Z. Jinxiang and J. Guanghui, 2014: Clinical characteristics of human infection with a novel avian-origin influenza A (H10N8) virus. Chinese Medical Journal,127, 3238-3242.
Zhang, Z., S. Hu, Z. Li, X. Wang, M. Liu, Z. Guo, S. Li, Y. Xiao, D. Bi and H. Jin, 2011: Multiple amino acid substitutions involved in enhanced pathogenicity of LPAI H9N2 in mice. Infection, Genetics and Evolution,11, 1790-1797.
Zhao, Z., C. Yi, L. Zhao, S. Wang, L. Zhou, Y. Hu, W. Zou, H. Chen and M. Jin, 2014: PB2-588I enhances 2009 H1N1 pandemic influenza virus virulence by increasing viral replication and exacerbating PB2 inhibition of beta interferon expression. Journal of Virology,88, 2260-2267.
Zheng, Y., C. Ye, L. Yunfei, X. Xiuhong, Q. Zhiping, D. Lowrie, L. Xinian, W. Yanbing, Z. Qi and L. Shuihua, 2013: Clinical comparison of one recovered case and one fatal case of human infection with H7N9 avian influenza in Shanghai Public Health Clinical Center in China. The Malaysian Journal of Medical Sciences,20, 76-79.
Zheng, Y.-F., Y. Cao, Y.-F. Lu, X.-H. Xi, Z.-P. Qian, L. Douglas, X.-n. Liu, Y.-b. Wang, Q. Zhang and S.-H. Lu, 2013: A case with non-typical clinical course of H7N9 avian influenza. Chinese Medical Journal,126, 4399-4399.
Zhou, P., W. Zhu, H. Gu, X. Fu, L. Wang, Y. Zheng, S. He, C. Ke, H. Wang and Z. Yuan, 2014: Avian influenza H9N2 seroprevalence among swine farm residents in China. Journal of Medical Virology,86, 597-600.
Zhu, G., R. Wang, F. Xuan, P. Daszak, S. J. Anthony, S. Zhang, L. Zhang and G. He, 2013: Characterization of recombinant H9N2 influenza viruses isolated from wild ducks in China. Veterinary Microbiology,166, 327-336.
Zhu, H., D. Wang, D. J. Kelvin, L. Li, Z. Zheng, S. W. Yoon, S. S. Wong, A. Farooqui, J. Wang and D. Banner, 2013: Infectivity, transmission, and pathology of human-isolated H7N9 influenza virus in ferrets and pigs. Science,341, 183-186.
Zhu, W., Y. Zhu, K. Qin, Z. Yu, R. Gao, H. Yu, J. Zhou and Y. Shu, 2012: Mutations in polymerase genes enhanced the virulence of 2009 pandemic H1N1 influenza virus in mice. Plos One,7,
Zhu, Y., S. Hu, T. Bai, L. Yang, X. Zhao, W. Zhu, Y. Huang, Z. Deng, H. Zhang and Z. Bai, 2014: Phylogenetic and antigenic characterization of reassortant H9N2 avian influenza viruses isolated from wild waterfowl in the East Dongting Lake wetland in 2011–2012. Virology Journal,11, 1-11.
Zmasek, C. M. and S. R. Eddy, 2001: ATV: display and manipulation of annotated phylogenetic trees. Bioinformatics,17, 383-384.
DISCLAIMER
This article was translated to enable Bahasa Indonesia readers to better understand the topic explained inside the article in their native language. The translator made no profit in translating the article.