

Oleh: Colin R. Parrish dan Edward J. Dubovi
Diterjemahkan oleh: Agna D. Lantria
DISCLAIMER
This article was translated to enable Bahasa Indonesia readers to better understand the topic explained inside the article in their native language. The translator made no profit in translating the article.
Virus canine Influenza H3N8
Kemunculan
Virus equine influenza (EIV) H3N8 telah bersirkulasi pada kuda sejak tahun 1963, ketika muncul pada kuda di Amerika Selatan, kemungkinan berasal dari penularan virus avian influenza [29]. Infeksi virus tersebut pada anjing telah diamati pada beberapa kejadian, seperti wabah kecil yang terjadi pada anjing foxhound di Inggris tahun 2022, dengan beberapa hewan juga terinfeksi selama tahun 2003 [8], sementara infeksi tunggal pada anjing yang berdekatan dengan kuda terinfeksi (10 dari 40 anjing diamati) terdeteksi pada saat berlangsungnya wabah virus EIV pada kuda di Australia tahun 2007 [20]. Tetapi, pada kasus-kasus ini virus mati setelah satu kali infeksi tunggal atau setelah beberapa bulan penularan. Sebaliknya, kemunculan virus canine influenza (CIV) H3N8 yang sukses pertama kali dikenalkan tahun 2004 saat wabah penyakit respirasi berat terlihat pada anjing greyhound yang berada dalam fasilitas pelatihan di negara bagian Florida, Amerika Serikat [7]. Sebelum dan selama tahun 2004 virus dibawa oleh anjing greyhound terinfeksi ke beberapa pacuan anjing di beberapa negara bagian Amerika Serikat, demikian juga anjing-anjing yang lain termasuk yang berada dalam shelter di beberapa negara bagian yang sama. Anjing-anjing yang terinfeksi virus CIV terutama menderita penyakit pernafasan atas yang menyebar cepat di dalam setiap fasilitas pemeliharaan, meskipun beberapa menunjukkan penyakit yang lebih berat termasuk bronkopneumonia hemoragika [7]. Virus influenza A H3N8 diisolasi dan menunjukkan hubungan kekerabatan dekat dengan virus EIV H3N8, dan lebih spesifiknya berasal dari strain EIV clade 2 Florida yang bersirkulasi pada kuda di Florida [7]. Analisis sera yang dikumpulkan dari anjing di Florida di tahun 1990an dan 2000an menunjukkan bahwa serum positif pertama kali dikoleksi tahun 2000, mengindikasikan bahwa virus mungkin ditularkan pada anjing tahun 1999 atau 2000, dan karena itulah telah bersirkulasi selama 3 atau 4 tahun sebelum pertama kali diidentifikasi sebagai penyebab penyakit. Uji serologis menunjukkan bahwa antibodi H3 dalam serum anjing yang dikumpulkan dari anjing greyhound yang terlibat pada fasilitas pelatihan dan pacuan yang berbeda, demikian juga shelter terdampak, memperlihatkan proporsi jumlah seropositif yang berbeda-beda (sampai 100%) dari anjing-anjing dalam setiap fasilitas [2, 7, 30].
Analisis sekuens virus menemukan bahwa wabah virus CIV H3N8 diinisiasi oleh satu virus yang ditularkan dari kuda, dan keturunan dari virus tersebut telah menyebar terus menerus di antara anjing [15, 32]. sejak saat itu (Gambar 22.1A dan C), tanpa adanya bukti reassortment dengan virus influenza A lainnya [15, 32]. Garis keturunan virus CIV pada anjing dengan cepat berpisah cabang dari virus EIV yang tetap bersirkulasi pada kuda, dan telah mendapatkan substitusi spesifik dalam setiap segmen gen untuk membentuk clade spesifik CIV untuk setiap segmen gen, yang sekarang berbeda dengan sekuens dari setiap virus yang bersirkulasi pada kuda. Meskipun ciri-ciri spesifik virus canine masih belum diidentifikasi dengan jelas, beberapa perubahan sekuens spesifik CIV mungkin telah terseleksi khusus untuk adaptasi pada anjing [15]. Perubahan di dalam dan di dekat tempat perlekatan reseptor protein HA1 menunjukkan kemampuan merubah perlekatan kepada glycan sulfat, dan mungkin menyebabkan perubahan lain terhadap perlekatan asam sialik [6], yang mungkin mendukung replikasi virus dalam saluran respirasi anjing. Strain virus EIV telah menunjukkan kemampuan menginfeksi anjing [45, 46], demikian juga kultur jaringan canine trachea [13], sehingga jumlah adaptasi yang diperlukan agar virus H3N8 mampu menginfeksi anjingnya tampaknya rendah, dan perubahan tambahan dapat mendukung penularan virus antar anjing. Saat ini belum ada bukti penularan virus CIV kembali ke kuda secara alami di lapangan, dan mungkin memang terdapat barrier untuk infeksi semacam itu, karena isolat CIV bereplikasi kurang efisien pada kuda yang diuji tantang eksperimental atau pada kultur sel trakea kuda [31, 44, 46]. Virus EIV H3N8 juga telah diisolasi dari babi yang menunjukkan gejala klinis di China [42], demikian juga dari unta Bactrian [47].
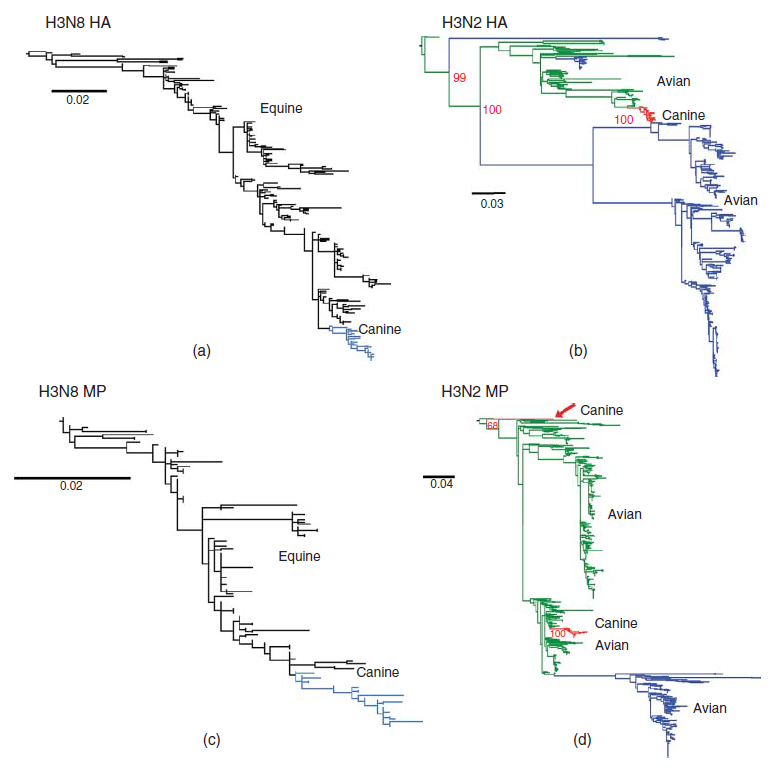
Gambar 22.1 Filogeni menunjukkan sumber awal virus CIV yang telah bersirkulasi secara luas, yaitu H3N8 yang muncul di Amerika Utara sekitar tahun 2000 sebagai varian virus EIV, dan virus H3N2 yang muncul di China dan Korea selama tahun 2005 sebagai varian virus avian. Sekuens HA CIV H3N8 (A) dan MP (C ), dibandingkan dengan sekuens virus EIV H3N8 dari virus-virus yang dikumpulkan dari kuda pada waktu yang berbeda-beda setelah virus muncul tahun 1963. Isolat virus-virus equine ditunjukkan dengan warna hitam, dan virus-virus canine ditunjukkan dengan warna biru. Sekuens HA H3N2 (B) dan MP (D) dibandingkan dengan sekuens virus avian influenza yang ada dalam database. Sekuens CIV ditunjukkan dengan warna merah, virus avian Eurasia ditunjukkan dengan warna hijau, dan virus-virus Amerika ditunjukkan dengan warna biru. Dimodifikasi dari gambar 1 dalam Hayward, J.J., E.J. Dubovi, J.M. Scarlett, S. Janeczko, E.C. Holmes, dan C.R. Parrish. 2010. Mikroevolusi virus canine influenza dalam shelters dan epidemiologi molekulernya dalam Amerika Serikat. Journal of Virology 84:12636-12645, dan Gambar 1 dari Zhu, H., J. Hughes, dan P.R. Murcia. 2015. Asal dan dinamika evolusi virus canine influenza H3N2. Journal of Virology 89:5406-5418.
Epidemiologi, persebaran dan pengendalian
Sejak 2004, virus CIV telah menyebar kepada setidaknya 11 negara bagian di Amerika Serikat, seringkali sebagai akibat dari transportasi anjing greyhound pacuan yang terinfeksi. Pada waktu yang sama virus CIV juga menginfeksi anjing peliharaan, demikian juga anjing dalam animal shelter dan kennel di beberapa wilayah Amerika Serikat [9, 30]. Tetapi, sejak 2007, virus CIV telah bertahan dalam sejumlah kecil fasilitas animal shelter berskala besar di kota-kota metropolitan besar yang memiliki cukup hewan rentan yang masuk secara reguler, demikian juga jumlah hewan yang menetap, yang memungkinkan transmisi dan maintenance virus berkelanjutan [9, 15]. Analisis sekuens virus menunjukkan bahwa populasi tersebut dapat mempertahankan virus-virus yang sama dalam sirkulasi berkelanjutan, najing-anjing terinfeksi dan virusnya sering dipindahkan dari shelter-shelter tersebut ke populasi anjing yang lain yang mengakibatkan wabah [30]. Wabah sekunder ini terutama terjadi shelter-shelter kecil, boarding kennel kecil, atau di antara anjing-anjing peliharaan, dan pada umumnya tidak berlangsung lama, karena besarnya populasi dan tingkat keluar masuknya anjing rentan tidak cukup untuk menyokong penularan berkelanjutan, karena itulah wabah menghilang dalam waktu beberapa minggu sampai bulan setelah memasuki shelter [2, 9, 30].
Penyakit
Virus CIV pada anjing seringkali menyebabkan penyakit pernapasan dengan gejala klinis ringan atau subklinis, meskipun penyakit berat kadangkala dihubungkan dengan infeksi. Penyakit ringan antara lain batuk yang biasanya lembab, meskipun bisa juga batuk kering, dan bisa jadi ada leleran hidung. Gejala dapat berlangsung 7 – 10 hari, dan akhirnya hewan sembuh dengan lancar. Anjing yang menunjukan penyakit berat mungkin mengalami demam tinggi (diatas 42o C) dan menunjukkan gejala klinis sangat cepat. Virus ditemukan di banyak jaringan pernafasan, dan infeksi trakea, bronki, dan paru-paru umumnya terlihat (Gambar 22.2) [4]. Pneumonia, termasuk pneumonia hemoragika juga dapat muncul.
Keparahan penyakit dapat bergantung kepada keadaan tertentu saat infeksi, dan gejala akan diperparah oleh adanya ko-infeksi dengan patogen lain. Inokulasi dengan virus EIV saja pada anjing akan berakibat infeksi subklinis, yang dapat merefleksikan tahapan awal wabah CIV pada anjing greyhound, yang tidak terdeteksi sebagai penyakit baru selama periode beberapa tahun. koinfeksi anjing oleh virus CIV dan patogen lain menghasilkan penyakit yang lebih berat, dan dalam salah satu studi ko-infeksi bersama Streptococcus equi subspecies zooepidemicus mengakibatkan penyakit yang jauh lebih berat dan menyerang paru-paru jika dibandingkan dengan infeksi tunggal dengan masing-masing patogen [21].
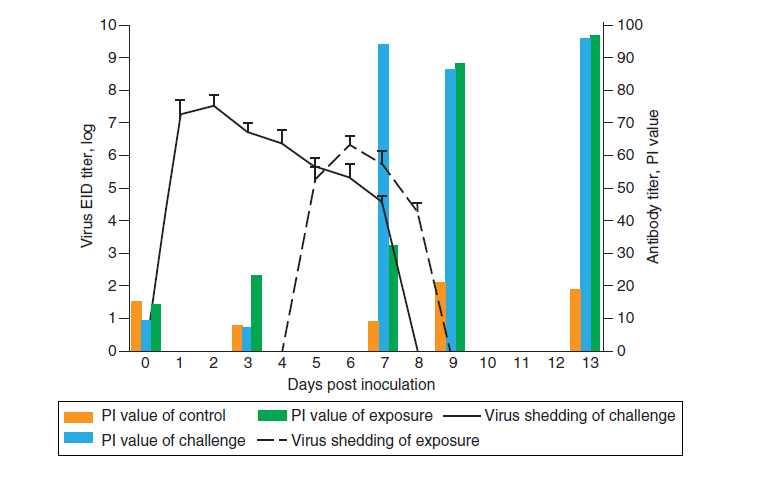
Gambar 22.2 Kurun waktu replikasi virus canine H3N2 pada anjing. Pendedahan virus dan respons serologis pada anjing beagle setelah eksperimen kontak penularan virus CIV H3N2 pada anjing. EID = dosis infeksius dalam telur; PI = persentase inhibisi. sumber : Gambar 1 dari Song, D., C. Lee, B. Kang, K. Jung, T. Oh, H. Kim, B. Park dan J. Oh. 2009. Experimental infection of dogs with avian-origin canine influenza A virus (H3N2). Emerging Infectious Diseases 15:56-58. Diberikan dengan izin.
Vaksinasi
Sejumlah vaksin telah dikembangkan untuk virus CIV H3N8, meliputi sejumlah vaksin virus inaktif (killed). Vaksin-vaksin ini meliputi sejumlah vaksin virus inaktif (killed), yang mengurangi gejala infeksi dan penyakit klinis jika diberikan dua dosis 3 minggu terpisah [10, 21]. Vaksin-vaksin ini umumnya direkomendasikan untuk anjing yang beresiko tinggi terhadap infeksi respiratori, tetapi saat ini tidak diberikan secara rutin kepada kebanyakan anjing peliharaan. Vaksin vektor canarypox eksperimental yang mengekspresikan protein HA dari virus EIV atau CIV menunjukkan efektivitas dalam mengurangi replikasi virus dan keparahan penyakit [18].
Canine Influenza H3N2
Kemunculan dan persebaran setelahnya
Penyakit respirasi anjing di China dan Korea dikenali disebabkan oleh virus influenza H3N2 selama tahun 2006 dan 2007, dan mungkin muncul melalui penularan langsung dari virus AI, kemungkinan dari virus yang bersirkulasi pada pasar unggas hidup (LBM) di Korea atau China [35]. Virus tersebut menyebar secara luas diantara anjing-anjing di Korea Selatan dan di beberapa wilayah China [25, 26], dan juga dihubungkan dengan wabah penyakit respirasi di Thailand tahun 2012 [3]. Studi serologis retrospektif mendemonstrasikan keberadaan antibodi anti-CIV dari anjing di Korea tahun 2005 [23]. Asal tepatnya virus CIV H3N2 ini masih belum diketahui, analisa filogeni sekuens virus menunjukkan bahwa virus baik dari China maupun Korea berhubungan dekat dengan leluhur keturunan canine (Gambar 22.1B dan D), mengindikasikan bahwa terdapat transfer virus yang cepat antara China dan Korea, dan melampaui jarak yang luas di dalam wilayah China sendiri [39, 43, 51], dan virus yang sama juga dihubungkan dengan wabah penyakit respirasi di Thailand tahun 2012 [3].
Infeksi pada hospes-hospes lain dan reassortment
Virus CIV H3N2 mampu untuk menginfeksi kucing dan telah menyebabkan wabah alami pada kucing dibawah kondisi tertentu [16]. Saat diuji kemampuan replikasinya pada hewan lain, ditemukan bahwa ayam, babi, mencit, babi guinea, dan musang yang diinokulasi menunjukkan penyakit yang tidak begitu jelas, meskipun serokonversi terlihat pada musang, babi guinea dan ayam, tapi tidak pada babi dan mencit [27]. Pendedahan virus dan lesi paru-paru terlihat pada babi guinea dan musang [27]. Eksperimen penularan dari anjing terinfeksi kepada kucing yang tinggal serumah juga dilaporkan, sementara musang menjadi terinfeksi setelah eksperimen inokulasi [19], dan beberapa penyebaran alami terbatas antar musang juga terdeteksi [24].
Virus hasil reasortan mengandung segmen virus H3N2 dan segmen gen lainnya dari virus manusia telah dideskripsikan, termasuk satu virus yang mengandung 7 segmen gen dari virus pandemi H1N1 dan segmen HA dari virus canine [38].
Penyakit
Virus H3N2 tampaknya secara umum menyebabkan penyakit saluran pernafasan atas, meskipun keparahan penyakit mungkin lebih besar daripada yang disebabkan H3N8. Dalam eksperimen infeksi, penyakit klinis terlihat 1 hari pasca inokulasi, dengan skor klinis tertinggi antara hari ke-4 dan 6, dengan sebagian besar replikasi terjadi pada saluran pernafasan, meskipun beberapa virus dideteksi di beberapa organ lain [36, 37, 48] (Gambar 22.3). Beberapa infeksi lebih berat juga dilaporkan, kemungkinan berhubungan dengan infeksi CIV dengan patogen respirasi lainnya, atau dengan ekspresi gen yang menginduksi inflamasi dan apoptosis [17].
Epidemiologi
Penyebaran virus CIV H3N2 tampaknya berbeda dengan yang terlihat pada virus H3N8 di Amerika Serikat, karena level infeksi yang tinggi dilaporkan dalam sampling serologi yang sudah dilaksanakan, yaitu level antibodi yang lebih tinggi pada anjing peliharaan rumahan, demikian juga pada anjing di beberapa shelter dan kennel yang memiliki populasi besar dan tingkat keluar masuk tinggi. Sebagian besar studi yang telah memeriksa antibodi H3N2 pada anjing memiliki berdasarkan kenyamanan pengambilan sampel, tetapi persentase anjing yang seropositif berkisar dari kurang dari 5% sampai 30% atau lebih pada anjing-anjing yang berasal dari beberapa wilayah [50], dan infeksi level tinggi telah dilaporkan pada anjing pertanian di China [40].
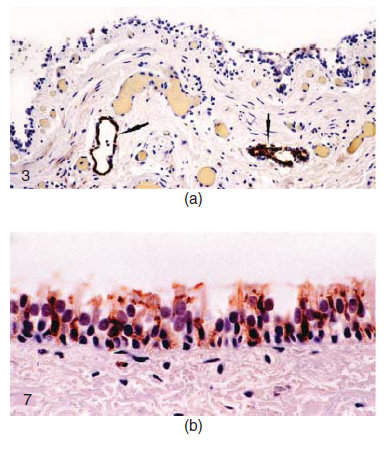
Gambar 22.3 Infeksi dan replikasi virus CIV H3N8 pada anjing setelah infeksi alami atau eksperimental. (A) Bronchus dari anjing greyhound yang diinfeksi spontan dengan virus canine influenza. Terdapat erosi epitel fokal pada epitel permukaan. Sitoplasma sel epitel dan sitoplasma makrofag luminal positif mengandung antigen hemagglutinin (tanda panah). Pewarnaan imunohistokimia adalah untuk antigen virus H3. (B) Trakea anjing 5 hari pasca inokulasi virus canine influenza. Antigen hemaglutinin virus terdapat dalam sitoplasma sel bersilia dan non-silia demikian juga sel basal. Imunohistokimia untuk antigen virus H3. Diambil dari Gambar 3 dan 7 dari Castleman, W. L., J. R. Powe, P. C. Crawford, E.P. Gibss, E.J Dubovi, R.O. Donis, dan D. Hanshaw. 2010. Canine H3N8 influenza virus infection in dogs and mice. Veterinary Pathology 47:505-517, dengan izin dari Sage.
Vaksinasi
Vaksin inaktif komersial telah dikembangkan untuk digunakan pada kucing di Amerika Serikat. Sifat dan penggunaanya serupa dengan vaksin H3N8 yang dijelaskan di atas. Vaksin eksperimental juga telah dideskripsikan [22].
Infeksi influenza lain pada anjing (dan kucing)
Sejumlah infeksi pada anjing oleh virus influenza lain yang bersirkulasi pada manusia dan unggas telah dilaporkan, dengan antibodi terdeteksi pada sebagian besar kasus, dan penyakit juga teramati pada beberapa kejadian. Banyak studi telah mendeteksi keberadaan virus pandemi influenza H1N1 (H1N1p) atau virus influenza manusia lainnya pada anjing dan kucing. Sekitar 0,7% sera positif untuk H1N1p dilaporkan pada anjing di Italia [11], dan virus ini juga dihubungkan dengan wabah penyakit klinis pada kucing, dengan seropositif sebesar 55%. Pengujian pada anjing dan kucing di Jepang menggunakan uji serologis menunjukkan bahwa 3,8% kucing dan 2,1% anjing positif antibodi H3 manusia [33], sementara jumlah yang lebih tinggi (20-50% seropositif dengan uji HAI) dilaporkan pada kucing dengan virus influenza manusia di Amerika Serikat [1] dan presentasi sera positif serupa juga dilaporkan pada anjing di Amerika Serikat [28, 34]. Alasan tingginya proporsi seropositif pada hewan di Amerika dibandingkan dengan negara lain belum diketahui, tapi mungkin berhubungan dengan pengujian yang digunakan untuk mendeteksi antibodi spesifik. Virus AI H5N2 diisolasi dari anjing di China tahun 2009 [49]. Studi serologis pada anjing liar yang mengunjungi pasar unggas hidup (LBM) di China menunjukkan persentase antibodi rendah namun konsisten terhadap infeksi H9N2, H3N2 dan H5N1 [41]. Anjing dan kucing mungkin juga dapat terinfeksi virus AI H5N1 setelah diuji tantang secara eksperimental [5], dan infeksi mungkin juga terjadi setelah memakan daging hewan yang terinfeksi.
Kesimpulan
Infeksi virus influenza A pada mamalia telah didokumentasikan dengan baik setidaknya dalam 100 tahun terakhir, dan wabah yang meluas dan epidemi yang berkelanjutan telah diamati pada kuda, anjing laut, babi, mink, dan manusia, tetapi sampai saat ini infeksi influenza pada anjing tidak secara luas dikenali [14]. Akan tetapi, selama 16 tahun terakhir, virus influenza A H3N2 dan H3N8 telah muncul dalam kejadian yang berbeda, dan telah menyebar menjadi epidemi dalam populasi anjing terpisah. Selain itu, beberapa wabah terbatas juga sudah dilaporkan pada anjing, dan juga terdapat infeksi spillover dari hospes anjing kepada hospes lain, termasuk kucing. Terdapat juga peningkatan bukti level infeksi rendah tapi konsisten pada anjing oleh beberapa virus influenza musiman manusia yang bersirkulasi, dan juga oleh virus avian, meskipun mungkin dengan gejala ringan atau tidak ada penyakit. Karena anjing seringkali terpapar virus dari hewan lain, tampanua terdapat barrier signifikan terhadap infeksi cross-spesies, tetapi barrier ini dapat diatasi oleh virus influenza dari tipe yang berbeda di bawah kondisi tertentu.
DISCLAIMER
Artikel terjemahan ini dimaksudkan bagi pembaca berbahasa Indonesia untuk dapat mempelajari artikel keilmuan terkait. Penerjemah tidak mendapatkan keuntungan apapun dari kegiatan penerjemahan ini.