Bab 7. Infeksi dan ekologi virus avian influenza pada unggas liar
Oleh David E. Stallknecht dan Justin D. Brown
Diterjemahkan oleh: Agna D. Lantria
DISCLAIMER
Artikel terjemahan ini dimaksudkan bagi pembaca berbahasa Indonesia untuk dapat mempelajari artikel keilmuan terkait. Penerjemah tidak mendapatkan keuntungan apapun dari kegiatan penerjemahan ini.
Lima puluh tahun telah berlalu setelah sejak virus avian influenza (AIV) pertama kali dilaporkan dari Common terns (Sterna hirundo) di Afrika Selatan [14]. Meskipun secara historis berskala terbatas, studi lapangan yang dilakukan dari tahun 1970-an sampai tahun 1990an sangat meberikan dampak peningkatan pemahaman tentang sejarah alami virus AI pada reservoir burung liar. Ketertarikan saintifik tentang topik ini meningkat pada tahun 2002, sebagai respons terdeteksinya virus AI sangat patogenik (HP) H5N1 pada burung liar di Hong Kong yang selanjutnya menyebar ke populasi unggas di Eurasia dan Afrika. Sebagai akibatnya, jumlah publikasi yang berhubungan dengan surveilans burung liar dan ekologinya meningkat secara dramatis [100]. Ledakan terbaru dalam surveilans dan aktivitas riset ini telah berakibat perspektif global yang lebih berkembang, dan juga pemahaman yang lebih mendetail tentang kisaran hospes, reservoir burung liar, keragaman genetic virus AI dalam populasinya, pola infeksi secara spatial dan temporal, mekanisme transmisi dan maintenans, dan resiko yang terkait dengan virus seperti HPAI H5N1 yang dapat memiliki kesamaan antara populasi unggas domestic dan populasi unggas liar. Tujuan Bab ini adalah untuk memberikan sebuah gambaran pemahaman kita yang terbaru tentang sejarah alami dan epidemiologi virus AI pada reservoir burung liar, dan untuk mengidentifikasi celah-celah dalam pemahaman ini yang perlu diatasi melalui penelitian di masa mendatang.
Ada beberapa review yang sudah dipublikasikan yang menyediakan informasi mendetail tentang kisaran hospes AIV pada unggas liar [157, 206, 211]. Dengan bertambahnya jumlah spesies akibat dari bertambahnya surveilans dan penelitian, daftar yang sudah diperbaharui disajikan dalam tabel 7.1. hasil yang disajikan dalam tabel ini masih tidak mewakili kisaran hospes yang sebenarnya, karena hanya berdasarkan laporan isolasi virus. Terdapat laporan tambahan hasil positif polymerase-chain reaction (PCR) dari spesies-spesies yang belum pernah diisolasi virus AI darinya, dan juga ada banyak sekali spesies yang belum pernah diujikan.
Kisaran hospes hanya setengahnya didefinisikan oleh kerentanan spesies. Alasan dari hal ini terkait dengan perbedaan genetic, geografi, dan tingkah laku yang dimiliki oleh lebih dari 9000 spesies burung liar yang ada di seluruh dunia [64]. Meskipun jumlah dan keragaman spesies burung dimana virus AI telah diisolasi sudah ekstensif (Tabel 7.1), ada beberapa karakterisitik taksonomi dan tingkah laku yang mendefinisikan sebagian besar spesies yang telah positif virus AI ini. sebagian bear berhubungan dengan habitat akuatik, dan sebagain besar spesies termasuk kedalam satau atau dua ordo Avian, Anseriformes (itik, angsa) dan Charadriiformes (burung camar, terns, dan burung pantai). Bahkan didalam ordo-ordo ini pun, tampak jelas adanya variasi dalam kemungkinan hasil tes positif virus AI. Dalam ordo Anseriformes misalnya, sebagian besar isolasi virus AI telah dilaporkan dari subfamily Anatinae (dabbling and diving ducks/ itik yang mencari makan di permukaan air dan itik yang mencari makan dengan menyelam ke dalam air). Variasi juga ditemukan pada Anatinae, dimana terdapat tingkat isolasi yang berbeda pada subfamily ini. Pada umumnya, lebih banyak isolasi yang dilaporkan dari itik yang mencari makan di permukaan air dibandingkan dengan itik yang mencari makan dengan menyelam dan angsa [82, 148, 206, 243], dan diantara itik yang mencari makan di permukaan, sebagian besar isolasi telah dilaporkan dari mallard (Anas platyrhynchos) [155].
Perbedaan ini utamanya merefleksikan perbedaan tingkah laku yang terkait dengan spesies (pilihan habitat, strategi mencari makan, dan pola migrasi) yang mempengaruhi transmisi, akan tetapi beberapa perbedaan ini bisa saja berhubungan dengan upaya sampling yang bisa saja bias terhadap spesies yang berjumlah banyak dan mudah di-sampling yang secara historis memiliki prevalensi tinggi terhadap virus AI.
Pola yang lebih restriktif lagi ditemukan pada Charadriiformes. Meskipun virus AI telah diisolasi dari spesies yang berasal dari setidaknya empat family (Scolopacidae, Charadriidae, Laridae, dan Alcidae), burung camar (Laridae) telah secara konsisten menjadi sumber hasil positif paling banyak. Sebaliknya, laporan isolasi virus dari unggas lain dalam family ini, seperti burung laut (tern), relatif sedikit. Variasi terkait speses juga tampak jelas pada burng-burung pantai (Scolopacidae dan Charadriidae), dan secara global, mengkonfirmasi bahwa infeksi virus AI hanya dilaporkan pada sejumlah spesies yang terbatas (sebagian bear termasuk dalam Scoopacidae), dan meskipun telah terdeteksi, prevalensinya seringkali sangat rendah [36, 61, 148].
Isolat juga telah dilaporkan dari spesies lain yang tinggal di habitat akuatik, meliputi unggas yang ada pada ordo Ciconiiformes, Gaviiformes, Gruiformes, Pelecaniformes, Podocipediformes, dan Procellariiformes. Secara kolektifnya, burung ini meliputi sedikit spesies positif (Tabel 7.1). Sangat mungkin bahwa banyak hasil positif ini berhubungan dengan tumpahan (spillover) virus dari itik dan burung camar dalam habitat akuatik bersama. Hal ini yang terjadi pada isolat-isolat yang baru-baru ini dilaporkan dari American white pelicans (Pelacanus occidentalis) [127] dan red-necked grebes (Podiceps grisegena) [130] yang diambil sampelnya dari habitat burung air di Minnesota. Pada kedua kasus, virus-virus yang diisolasi dari spesies-spesies ini jelas merefleksikan asal dari itik ataupun asal campuran dari itik dan burung camar.
Terdapat laporan virus AI dari spesies dalam ordo Columbiformes, Piciformes, dan Passeriformes yang umumnya dihubungkan dengan habitat terrestrial, akan tetapi ada beberapa isolasi yang dilaporkan. Pada ulasan yang baru tentang virus AI pada passerine [200] yang mencakup hasil laporan positif dari isolasi virus, PCR dan uji serologis, telah disimpulkan bahwa hanya sedikit bukti yang menunjukkan bahwa burung-burung ini terlibat dalam maintenans dan transmisi virus AI dibawah kondisi alami. Akan tetapi, dibawah kondisi dimana burung peridomestik berkontak dengan burung domestic yang terinfeksi virus AI, brung-burung darat ini bisa saja menjadi terinfeksi dan bisa saja memainkan peranan terbatas dalam transimisi [200].
Hubungan hospes ini telah disokong oleh studi serologis berskala-besar yang diselenggarakan baru-baru ini [24]. Meskipun begitu, pengujian serologis juga telah menyediakan wawasan dan pertanyaan terkait determinan kisaran hospes yang hanya berdasarkan deteksi virus saja. Sebagai contoh, hasil uji serologis memberikan bukti pertama bahwa itik terlibat dalam epidemiologi virus AI [9]. Penelitian yang lebih baru telah menunjukkan bahwa prevalensi antibody bisa jadi tinggi pada beberapa spesies burung dimana isolasi virus jarang dilaporkan atau dimana prevalensi infeksi biasanya berjumlah rendah secara konsisten. Hal ini telah terobservasi pada burung red knot (Calidris canutus) di Teluk Delaware [145], Canada goose (Branta canadensis) [82, 115], dan pink-footed goose (Anser brachyrhyncus) [101]. Hasil yang kontras ini menunjukkan bahwa banyak dari spesies-spesies ini terinfeksi selama hidup mereka, akan tetapi mereka tidak berkontribusi kepada kejadian prevalensi tinggi setiap tahunnya yang terdeteksi pada itik selama akhir musim panas dan musim gugur atau pada burung-burung pantai di Teluk Delaware selama musim semi. Karena banyak dari spesies ini memiliki hidup yang panjang, hal ini bisa saja terkait dengan struktur umur dalam populasi dan imunitas populasi yang dihasilkan darinya. Hal ini juga bisa saja berhubungan dengan tingkah laku (misalnya perilaku makan seperti digembalakan) dan juga berkurangnya waktu shedding virus.
Kerentanan spesies telah dievaluasi dalam penelitian eksperimental, akan tetapi informasi yang resedia masih terbatas. Secara eksperimental dimungkinkan untuk menginfeksikan AIV kepada hewan-hewan dalam grup taksonomi yang luas, akan tetapi perbedaan terkait spesies dalam kemampuan menginfeksi, durasi shedding virus, dan terdapat rute shedding dominan untuk setiap strain individual virus AIV [4, 5, 26, 97, 110, 196, 246], spesies unggas [35, 151], dan umur [33]. Studi eksperimental yang ada saat ini disokong oleh banyaknya laporan isolasi virus AI dari spesies sampai kelompok taksonomi, seperti Passeriformes, Psittaciformes, dan Galliformes. Meskipun begitu, pada hampir sebagian besar kasus ini unggas yang positif virus AI memiliki kontak langsung ataupun kontak potensial dengan unggas yang terinfeksi [3, 4, 193, 200]. Situasi yang serupa juga telah dilaporkan tentang virus HPAI H5N1 dibawah kondisi eksperimental [167, 169]. Pengakuan bahwa virus-virus ini memiliki potensi untuk menginfeksi spesies unggas yang beragam sangat penting dari sudut pandang pencegahan pemasukan virus ke dalam populasi hewan domestic, terutama berdasarkan fakta bahwa transmisi potensial mungkin tidak melibatkan secara langsung spesies yang mewakili hospes atau reservoir virus AI yang sudah dikenal dibawah kondisi alamiah. Pada saat ini variasi respons spesies terhadap infeksi virus AI belum dimengerti secara jelas, akan tetapi tampaknya tidak secara langsung berhubungan dengan distribusi reseptor asam sialic [56].
Telah ditetapkan bahwa semua AIV yang menginfeksi hospes unggas dan mamalia dalam sejarahnya berasal dari reservoir unggas liar. Pengecualian paling baru datang dari virus influenza kelelawar (H17 dan H18N11) yang baru-baru ini telah dilaporkan, akan tetapi sampai saat ini virus-virus ini belum diasosiasikan dengan unggas ataupun dengan tumpahan (spillover) ke hospes-hospes lain [223, 224]. Dalam mendefinisikan reservoir AIV penting untuk dimengerti bahwa hal ini mewakili sistem multispesies yang menyediakan lingkungan yang tepat untuk mempertahankan keberlangsungan hidup virus. Sangat penting pula untuk mengenali bahwa reservoir dapat berubah-ubah seiring dengan perpindahan virus masuk dan berevolusi di dalam populasi hospes yang baru. Sebuah reservoir didefinisikan sebagai “setiap benda hidup atau benda mati atau kombinasi dari keduanya yang berfungsi sebagai habitat pathogen yang mereproduksikan dirinya sedemikian rupa agar dapat bertransmisi kepada hospes yang sesuai” [233]. Seperti yang telah disebutkan sebelumnya, informasi kolektif tentang virus AI pada burung liar secara jelas mengidentifikasi dua grup taksonomi luas yang merepresentasikan tumpeng tindih dan pada beberapa kasus reservoir uni virus AI, yaitu Anseriformes dan Charadriiformes. Hal ini tidak mengatakan bahwa semua spesies di dalam kelompok ini berkontribusi sama rata dalam maintenans virus. Pada beberapa kasus, spesies individual dapat berkontribusi terhadap maintenans virus dalam bentuk yang untik. Sebagai contoh, meskipun itik Mallard adalah komponen penting terhadap reservoir virus AI pada itik liar dan seringkali diidentifikasi sebagai spesies paling penting yang berhubungan dengan maintenans virus AI, spesies ini seringnya hidup secara berdampingan dengan banyak spesies itik yang juga hidup dan mencari makan di permukaan air. Tambahan lagi, pada banyak lokasi wintering di Amerika Utara, seperti rawa-rawa pantai yang mengelilingi Semenanjung Meksiko, itik Mallard tidak mewakili spesies itik terbanyak, dan spesies lain, seperti blue-winged teal (Anas discors) atau green winged-teal (Anas crecca), bisa saja mewakili spesies utama dimana virus-virus ini dipertahankan secara musiman [207]. Spesies seperti northern shoveler (Anas clypeata), yang memiliki tingkah laku makan yang untuk, juga mungkin memberikan kontribusi unik dalam maintenans virus berdasarkan strategi makan yang meningkatkan kontak dengan sedimen yang berpotensi terkontaminasi virus AI [85]. Tambahan lagi, beberapa studi telah memberikan saran bahwa maintenans virus-virus ini pada itik mungkin tergantung pada interaksi antar subpopulasi burung residen, burung local, dan burung migran jarak jauh meskipun di dalam spesies yang sama [52, 86, 87, 207, 232, 235].
Terkait Charadriiformes, virus AI yang paling banyak diisolasi telah dilaporkan dari spesies dari dua family, Scolopacidae dan Laridae. Dalam family Laridae, terdapat bukti bahwa dua subtipe virus AI (H13 dan H16) di maintenans pada populasi burung camar [8, 51, 91]. Virus-virus ini hanya kadang-kadang saja dilaporkan dari itik dan burung wader, akan tetapi sangat prevalen pada populasi burung camar dalam koloni berbiak [234]. Virus H13 dapat diisolasi dari burung camar dengan prevalensi rendah sepanjang tahun, yang memebrikan saran bahwa virus ini dipertahankan setiap tahun di dalam populasi ini [69, 134]. Hal ini belum didemonstrasikan untuk virus H16 akan tetapi data untuk subtipe ini saat ini masih sedikit. Sebaliknya di dalam family Scilopacidae, studi molekuler yang melibatkan banyak gen virus AI belum menunjukkan perbedaan secara genetic antara virus AI burung-burung pantai dan itik [202]. Tambahan lagi, prevalensi infeksi yang sangat rendah telah didokumentasikan secara global dari spesies-spesies dalam famili ini [38, 61], dan sebagian besar isolasi virus AI di seluruh dunia telah dihubungkan pada satu spesies, burung ruddy turnstone (Arenaria interpres), pada satu lokasi (Teluk Delaware) dan pada satu waktu dalam satu tahun (Mei dan Juni) [145, 213]. Berdasarkan hasil surveilans ini saja, sangat sulit untuk menentukan apakah kelompok ini (Scolopacidae), atau bahkan burung ruddy turnstone, merupakan contributor signifikan terhadap keseluruhan reservoir virus AI, dan mungkin isolasi dari kelompok ini dapat merepresentasikan hospes tumpahan (spillover) terlokalisasi untuk virus-virus yang dimaintenans pada itik dan burung camar [145]. Sebaliknya, amplifikasi virus AI setiap tahunnya pada burung ruddy turnstone di Teluk Delaware bisa saja merepresentasikan komponen penting dalam maintenans dan pergerakan virus-virus ini ke arah utara semala migrasi musim semi [119].
Seperti telah dinyatakan sebelumnya, kisaran hospes hanya sebagiannya didefinisikan oleh kerentanan hospes. Begitu juga dokumentasi baik infeksi virus AI ataupun kerentanan terhadap virus tidak menentukan apakah satu spesies bersifat penting sebagai reservoir. Pada saat ini tampaknya terdapat sejumlah spesies terbatas yang berkontribusi terhadap maintenans dan transmisi virus-virus ini, akan tetapi hal ini dapat berubah dengan cepat jika terdapat perubahan yang memicu potensi maintenans dan transmisi.
Variasi spasial dan temporal pada prevalensi relative konsistentik, burung camar, dan burung-burung pantai berdasarkan benua, akan tetapi polanya dapat bervariasi secara global. Variasi ini dapat diatribusikan kepada perbedaan yang berhubungan dengan wilayah berbiak dan wilayah wintering (habitat/ tempat burung berada untuk menghindari musim dingin) dan keberadaan pemicu lingkungan yang berbeda-beda yang mempengaruhi distribusi burung. Pada itik Amerika Utara, prevalensi virus AI mencapai puncak pada akhir musim panas dan awal musim gugur, dan berhubungan dengan konsentrasi burung baru menetas yang bersifat rentan virus AI sebelum periode pre-migrasi [93]. Selama waktu-waktu ini, infeksi virus AI dapat melebihi 30% pada kelompok umur ini, dan sebagai konsekuensinya surveilans virus AI bisa sangat dipacu oleh konsentrasi burung-burung muda [243]. Pola temporal terobservasi pada itik berkorespondensi dengan pola spasial yang konsisten, dengan prevalensi virus AI yang tertinggi di Amerika Utara terobservasi pada unggas air pada wilayah berbiak dan wilayah staging (tempat berhenti sementara dalam perjalanan saat migrasi) di Canada dan Amerika Serikat bagian Utara. Selama burung bermigrasi ke arah selatan, prevalensi virus AI berkurang dengan cepat, dan pada wilayah wintering seringnya lebih rendah dari 1-2% [206]. Di Eropa dan Asia pola serupa juga terlihat, akan tetapi pada beberapa bagian eropa utara periode prevalensi tinggi tampaknya lebih Panjang sampai akhir musim gugur [238]. Alasan dari hal ini belum dimengerti, akan teteapi bisa saja berhubungan dengan perbedaan pilihan waktu migrasi atau kepada lokasi wilayah wintering yang lebih ke utara (bebas es). Seperti di Amerika Utara, prevalensi virus AI untuk wintering itik di Italia [39, 40], Perancis [126], Spanyol [166], Portugal [220], dan Georgia [134] umumnya rendah. Puncak prevalensi yang berhubungan dengan migrasi pada musim semi telah dilaporkan pada itik di Eropa dan Amerika Utara [81, 237], akan tetapi prevalensinya (sekitar 5-10%) relative rendah jika dibandingkan dengan migrasi pada musim gugur. Pola temporal kurang begitujelas untuk itik di Amerika Selatan, Afrika dan Australia [59, 62, 140, 156, 165]. Telah disarankan bahwa siklus influenza di Afrika adalah akibat dari pola musim hujan [37], dan bahwa musiman ini bisa jadi tidak begitu ekstrim dikarenakan tingkat rekrutmen burung-murung muda yang berangsur-angsur sebagai akibat dari perpanjangan musim berbiak [62]. Tidak seperti Afrika, yang merupakan rumah bagi burung residen yang tidak bermigrasi dan burung migrasi, populasi itik di Amerika Utara dan Australia hanya ada sedikit tumpang tindih atau tidak ada tumpang tindih sama sekali antara burung yang bermigrasi dari garis lintang utara dan spesies residen yang tidak bermigrasi [177, 227]. Pola infeksi temporal dan spasial pada unggas air lainnya, seperti angsa, kurang sekali deskripsinya, sebagiannya karena hasil isolasi yang kurang konsisten. Meskipun begitu, pada greater white-fronted geese (Anser albifrons) di Eropa, shedding virus yang berdurasi pendek, terutama dari traktus respiratori, terus terdeteksi secara konsisten selama musim dingin [116].
Pada burung camar, telah dilaporkan koneksi yang jelas antara prevalensi virus H13 dan H16 dan musim berbiak, dan seperti pada itik, puncaknya diasosiasikan dengan rekrutmen burung-burung muda (juvenile birds) [134, 233]. Untuk burung camar, hubungan temporal dengan skala yang lebih halus telah dideskripsikan termasuk infeksi virus AI yang berhubungan dengan kelas spesifik burung berumur muda pada sebuah koloni berbiak, dan perbedaan pemilihan waktu prevalensi puncak yang berhubungan dengan virus H13 dan H16 [234]. Diluar periode berbiak, virus Ai termasuk virus H13 dan H16 bisa saja diisolasi, akan tetapi prevalensinya sangat rendah [69].
Berdasarkan hasil yang utamanya berasal dari Teluk Delaware, puncak prevalensi virus AI utama pada Charadriiformes dilaporkan terjadi pada musim semi, dengan puncak yang lebih rendah terjadi pada musim gugur [110]. Meskipun hal ini terjadi pada burung camar, pola musiman dan spasial virus AI pada family-famili lain dibawa ordo yang sama belum dideskripsikan, dan pada umumnya prevalensi infeksi secara global rendah secara konsisten [38, 53, 61, 80, 210]. Untuk Teluk Delaware yang merupakan satu-satunya tempat di dunia dimana isolasi virus AI secara konstan telah dilaporkan, alasan tingginya prevalensi virus AI pada burung ruddy turnstones yang bermigrasi pada musim semi masih belum dimengerti [80, 110, 119, 121].
Pemahaman tentang reservoir unggas untuk AIV dan pola infeksi spasio-temporal pada hospes-hospes ini hanya dapat dimengerti dengan beberapa pengetahuan tentang imunitas komunitas. Hal ini terutama sangat penting pada populasi unggas liar, karena banyak dari spesies-spesiesnya hidup dalam waktu lama dan pada beberapa kasus mengalami tantangan berulang dari subtipe-subtipe AIV yang sangat beragam.
Saat ini hanya terdapat informasi terbatas tentang respons imun primer pada burung liar, dan karenanya pemahaman kita tentang perlindungan imun jangka panjang ataupun tentang respons terhadap infeksi yang banyak dan berkali-kali masih belum lengkap. Berdasarkan data lapangan, imunitas populasi tampaknya merupakan elemen yang sangat penting dalam siklus tahunan. Contohnya, telah diterima sejak lama bahwa prevalensi AIV yang tinggi pada itik muda selama tahap pre-migrasi berhubungan dengan konsentrasi burung yang belum memiliki kekebalan terhadap virus (naïve birds) [92]. Hubungan yang serupa juga tampaknya tampaknya terdapat pada burung camar yang terinfeksi virus LPAI H13 dan H16 pada koloni berbiak [234]. Pada burung-burung pantai di Teluk Delaware, prevalensi virus AI yang terobservasi pada level spesies berkorelasi dengan prevalensi antibodi yang telah ada sebelumnya pada burung ruddy turnstones dan red knot [145]. Durasi shedding virus pada itik Mallard yang terinfeksi secara alami menunjukkan bahwa shedding virus berkurang seiring waktu, memberikan saran bahwa imunitas yang didapat (acquired immunity) mengurangi shedding virus [122].
Studi eksperimental telah mendemonstrasikan efek potensial dari imunitas yang telah ada sebelumnya terhadap hasil akhir penyakit klinis dan shedding virus menggunakan virus LPAI dan HPAI [34, 36, 51, 109]. Berdasarkan sejarah re-isolasi dari sentinel atau burung yang ditangkap kembali, hasil yang serupa juga ditemukan pada studi lapangan [123, 221]. Temuan-temuan studi ini menyokong perkembangan imunitas lengkap pada uji tantang homolog atau imunitas parsial untuk uji tantang heterolog. Meskipun studi-studi ini utamanya masih terbatas pada itik Mallard, hasil yang sama telah dilaporkan pada Canada Geese [15]. Eksperimen uji tantang yang terkontrol, yang merefleksikan paparan jangka panjang dan infeksi ganda, seperti yang terjadi pada kondisi lapangan, saat ini sangat kurang. Dalam sudut pandang panjangnya umur spesies unggas dan adanya paparan virus AI kepada spesies-spesies ini setiap tahunnya atau bahkan sepanjang tahun, efek dan signifikansi potensial kejadian ini terhadap imunitas populasi merepresentasikan salah satu dari area studi paling penting yang masih belum banyak diteliti yang berhubungan dengan maintenans dan transmisi virus AI.
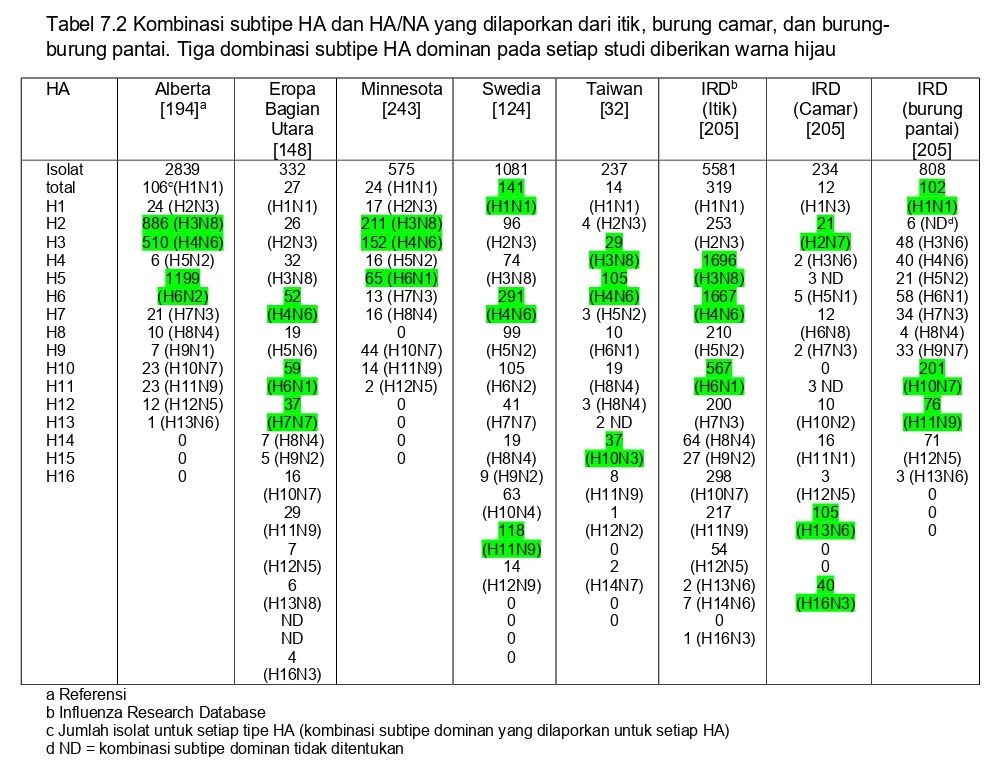
Terdapat 16 subtipe hemaglutinin (HA) dan 9 subtipe neuraminidase (NA) dari virus AI, dan sebagian besar kombinasinya telah dilaporkan dari burung liar [158]. Akan tetapi, subtipe-subtipe ini tidak terepresentasi secara seimbang diantara populasi burung liar dan variasi antar hospes, lokasi, musim, dan tahun juga dapat terjadi. Pada itik Amerika Utara, subtipe H3, H4, dan H6 mewakili subtipe HA predominan yang dilaporkan [119, 194, 206]. Beberapa subtipe HA (missal H11) sering diisolasi, dan lainnya (misal H8 dan H9) tidak umum [199, 206]. Trend-trend predominan ini ditemukan di Eurasia dan Amerika Utara [32, 124, 148, 243] (Table 7.2). Pada burung camar, keragaman subtipe tidak terdefinisikan secara jelas, akan tetapi virus H13 dan H16 secara konsisten mendominasi di seluruh dunia [8, 110, 119, 157]. Subtipe-subtipe lain, meliputi H1, H6, H10, dan H11 secara rutin diisolasi dari burung camar akan tetapi dengan prevalensi yang relative rendah [8, 69]. Sedangkan pada burung-burung pantai, pengetahuan kita tentang keragaman subtipe masih terbatas skop nya, dengan sebagian besar isolate berasan dari burung ruddy turnstone di Teluk Delaware, Amerika Serikat. Sembilan subtipe HA dari virus AI telah dilaporkan terjadi lebih sering pada Charadriiformes daripada pada itik, termasuk diantaranya virus AI H5, H7, dan H9 [119]. Pada Teluk Delaware, virus H6, H7, H10, H11, dan H12 sangat terwakili (over-represented) pada burung-burung pantai, virus-virus ini dapat saja muncul hampir setiap tahun, atau merupakan subtipe dominan pada satu tahun tertentu [213]. Seperti pada burung camar, subtipe H1, H6, H10 dan H11 terwakili dengan baik, maka subtipe H13 dan H16 sebaliknya malah tidak terwakili dengan baik (Tabel 7.2).
Efek lokasi dan musim terhadap keragaman subtipe belum cukup diselidiki, akan tetapi musim tampaknya lebih merupakan faktor yang lebih penting dibandingkan dengan lokasi. Pada itik Amerika Utara, virus H3, H4 dan H6 umum ditemukan selama periode staging dan migrasi, akan tetapi mereka sangat kurang terwakili saat migrasi musim semi, saat subtipe H7 dan H10 mendominasi [81, 178]. Karena prevalensi umumnya rendah di luar wilayah Teluk Delaware, variasi musiman dalam keragaman subtipe belum cukup diselidiki pada burung-burung pantai. Pada burung camar, hanya tampak hubungan antara virus H13 dan H16 dengan musim berbiak pada musim semi/panas.
Keragaman subtipe dapat bervariasi dari satu tahun ke tahun lainnya. Pada itik yang diambil sampelnya pada musim gugur, predominansi subtipe bergeser setiap tahunnya antara virus AI H3 dan H4 [119, 194, 243]. Alasan keadaan ini belum dimengerti, akan tetapi bisa saja berhubungan dengan imunitas populasi. Variasi tahunan dalam jumlah paling besar telah dilaporkan pada burung-burung pantai dari Teluk Delaware, dimana subtipe HA predominan antara tahun 2000 dan 2009 telah meliputi H1, H3, H4, H7, H9, H10, dan H12 [119, 213]. Variasi ini bisa saja merupakan akibat dari introduksi virus secara terbatas pada satu tahun tertentu, bersamaan dengan imunitas populasi. Pada beberapa tahun tertentu keragaman subtipe keseluruhan pada Teluk Delaware sangat rendah, dengan hanya beberapa subtipe yang terwakili [13]. Dengan pengecualian infeksi tahunan virus H13 dan H16 pada burung camar, hanya ada sedikit bukti adanya atau tidak adanya pola keragaman subtipe tahunan.
Belum dimengerti secara jelas mengapa dan bagaimana keragaman subtipe yang sedemikian rupa dipertahankan dalam populasi burung liar, atau mengapa beberapa subtipe HA mendominasi pada populasi burung tertentu ataupun pada musim tertentu. Ditambah lagi, saat ini kita masih tidak mengetahui mengapa beberapa kombinasi subtipe tertentu (H1N1, H2N3, H3N8, H4N6, H5N2, H6N1, H7N3, H8N4, H9N2, H10N7, H11N9, H12N5, H13N6, dan H16N3) seringkali terlalu terwakilkan dalam laporan isolate yang diambil dari itik. Hal ini terobservasi secara konsisten pada sampel yang diambil dari itik dari Eurasia dan Amerika Utara (Table 7.2). Pola-pola ini bisa saja berhubungan dengan keuntungan structural yang berhubungan dengan kebugaran keseluruhan dari kombinasi HA/NA tertentu, imunitas populasi, ataupun keduanya. Pola-pola kombinasi subtipe yang sama ini juga tampak pada virus AI yang diisolasi dari burung-burung pantai (predominannya merupakan isolate dari Teluk Delaware) (Tabel 7.2), akan tetapi mungkin karena keterbatasan isolat, virus H13 dan H16 tidak konsisten pada isolate yang berasal dari burung camar (Tabel 7.2).
Keragaman populasi virus AI dipertahankan pada burung-burung liar. Sebaliknya dari yang dipercaya sebelumnya bahwa virus-virus ini adalah dalam keadaan evolusi yang statis/tetap [68], studi yang dilakukan baru-baru ini telah mengungkapkan gambaran yang lebih kompleks dengan keragaman genetic berderajat tinggi dan sirkulasi beberapa jalur keturunan genetic secara bersamaan didalam satu subtipe [202]. Virus-virus AI bebas melakukan reassortasi di dalam tubuh reservoir burung liarnya [46, 83, 102, 245], dan studi genetic telah secara jelas mendemonstrasikan galur keturunan atau subgalur keturunan Amerika Utara, Eurasia dan Amerika selatan bagian selatan yang berbeda [43, 67, 165, 192, 202]. Perbedaan geografi dengan skala luas ini memberikan dugaan kemungkinan adanya isolasi global dengan percampuran virus yang jarang terjadi [141, 202]. Pola ini agak tidak terduga jika melihat tingkah laku migrasi pada banyak spesies aves yang dapat terinfeksi oleh virus AI yang setiap tahunnya bergerak melintasi benua-benua ini.
Keberadaan variasi gen virus AI Eurasia pada isolate Amerika Utara berasal dari itik, burung camar dan burung pantai telah dilaporkan, dan area telah diidentifikasi di Pasifik utara dan Atlantik utara dimana percampuran virus AI Eurasia dan Amerika Utara lebih mungkin terjadi [47, 58, 74, 118, 132, 164, 173, 174, 244]. Meskipun studi-studi ini menyokong aliran gen antar benua, tidak ada kejadian sampai saat ini dimana virus AI yang benar-benar ‘asing’ yang terdeteksi pada populasi-populasi ini. Dan meskipun diintroduksikan, kejadian transmisi seperti itu mungkin memiliki persistensi yang terbatas dalam populasi baru ini [179]. Tambahan lagi, hanya ada sedikit indikasi gen eksotik individu yang bertahan pada populasi “baru” setelah mereka diperkenalkan [120]. Salah satu contoh penting yang berhubungan dengan gen H6 Eurasia yang telah menggantikan virus H6 original dari Amerika Utara [12]. Hal ini tampaknya merupakan proses yang terjadi dalam jangka Panjang, dan mekanisme yang mendasari terjadinya pergantian ini dan potensi konsekuensi perubahan ini belum ditentukan. Sebuah contoh baru yang menggambarkan gen virus “baru” yang masuk ke dalam gen pool virus AI di Amerika utara melibatkan virus H14. Virus H14 pertama dideteksi di Amerika Serikat di tahun 2010, dimana pada waktu itu ia merupakan campuran dari garis keturunan Eurasia dan Amerika Utara. Proses sekuensing isolat berikutnya mengungkap reasortansi ekstensif dengan gen dari Amerika Utara, dan hanya gen HA H14 saja yang tetap tertinggal [17, 57, 176]. Introduksi sukses gen HA H10 dari Amerika Utara ke Australia juga telah dilaporkan [236].
Dari kumpulan literatur tampaknya pergerakan gen virus AI antar benua benar terjadi, dan meskipun jarang, pergerakan itu bisa saja berhasil. Sebaliknya, dengan kemungkinan pengecualian kejadian baru-baru ini yang berhubungan dengan virus HPAI H5N8, kurang adanya bukti perkenalan virus Eurasia dan Amerika Utara melewati benua melalui migrasi burung-burung liar.
Transmisi AIV dalam populasi burung liar utamanya terjadi melalui rute fekal atau oral [89, 190, 195]. Pada itik, replikasi terjadi terutama di saluran pencernaan [197], dan konsentrasi tinggi virus yang infeksius disebarkan melalui feses [91, 241]. Telah diduga bahwa air atau sedimen yang terkontaminasi mungkin merupakan sumber infeksi. Pandangan ini disokong oleh kisaran hospes yang meliputi banyak sekali burung-burung air, dan isolasi virus-virus ini dari air permukaan yang dipergunakan oleh burung-burung ini [75, 90, 108, 213]. Sebagai tambahannya, itik mallard bisa diinfeksi melalui berbagai rute (intranasal, intratrakeal, intraocular, intrakolakal, dan intra-ingluvial) yang dapat melibatkan kontak dengan air yang bersifat infektif [55]. Air permukaan dan air bawah tanah yang terinfeksi telah dianggap sebagai sumber virus AI jangka pendek dan jangka panjang yang memasukkan virus kedalam populasi unggas domestic [76].
Mekanisme transmisi yang berhubungan dengan kelompok unggas lainnya, seperti burung-burung pantai dan burung camar, masih kurang dimengerti, akan teteapi seperti pada itik, shedding virus berhubungan dengan baik rute kloaka maupun rute oroparing, dan virus dapat diisolasi dari feses.
Faktor hospes yang mempengaruhi kerentanan dan shedding virus
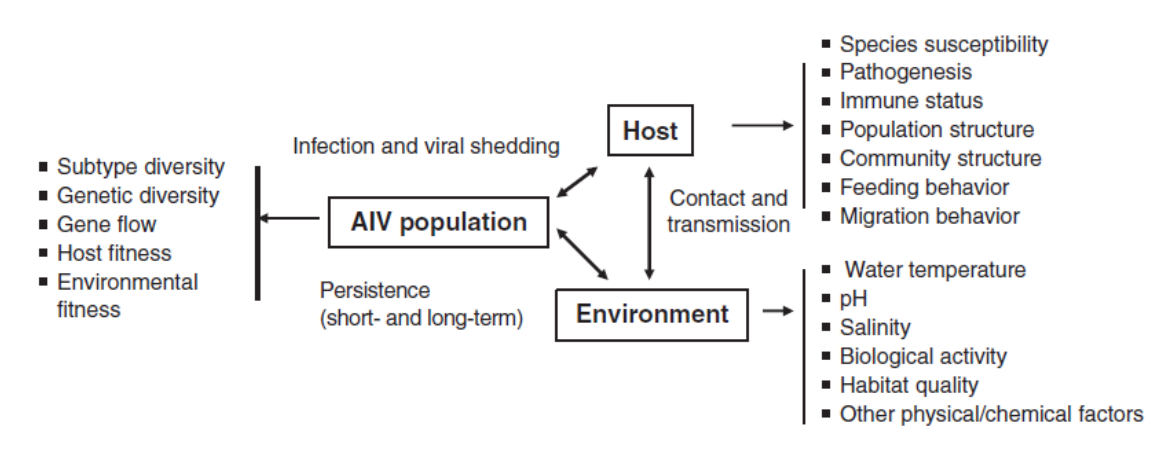
Ada banyak faktor hospes yang membawa potensi infeksi, shedding virus dan kontak dengan lingkungan (Gambar 7.1). Seperti yang telah disebutkan, banyak spesies burung liar rentan terhadap AIV, akan tetapi rute shedding virus durasi shedding, dan jumlah virus infeksius yang diekskresikan bervariasi antar spesies. Pada itik, shedding virus dapat diperpanjang sampai lebih dari 28 hari telah dilaporkan [44]. Meskipun perpanjangan shedding itu bisa saja terjadi, biasanya merupakan pengecualian, dan infeksi eksperimental pada itik mallard menunjukkan durasi shedding umumnya kurang dari 14 hari, dengan virus yang paling banyak diekskresikan pada hari ke-2 sampai ke-6 setelah infeksi [27, 33, 84]. Akan tetapi, di lapangan, estimasi itu mungkin tidak dapat diaplikasikan kepada burung dimana umumnya ia memiliki imunitas dari infeksi yang berulang. Hal ini dapat menjelaskan secara sebagian mengapa estimasi yang berdasarkan pada data lapangan menunjukkan periode shedding virus yang lebih pendek untuk itik Mallard, dengan estimasi minimum dan maksimum berkisar dari 3,1 sampai 8,3 hari [122]. Efek terhadap kerentanan dan shedding virus ini berpotensi meluas sampai kepada level populasi, terutama pada kaitannya dengan spesies yang memiliki hidup yang panjang, seperti angsa. Dalam kasus burung Canada geese, hanya sedikit burung terinfeksi yang terdeteksi, akan tetapi prevalensi antibodinya umumnya tinggi [115]. Hal ini menunjukkan bahwa meskipun sebagian besar burung terinfeksi sepanjang hidup mereka, mereka mungkin hanya sedikit berkontribusi terhadap transmisi tahunan.
Faktor lingkungan yang mempengaruhi infektivitas virus
Pentingnya persistensi AIV di lingkungan dalam transmisi dan dalam mempertahankan keberadaan virus masih belum jelas [106, 187, 212]. Investigasi awal persistensi lingkungan virus AI [242] mendemonstrasikan bahwa virus AI dapat tetap bersifat infektif di dalam feses atau air dalam waktu yang panjang. Penelitian selanjutnya telah mengkonfirmasi temuan ini pada banyak strain dan subtipe virus AI lainnya [20, 25, 138, 208, 209], dan juga mendemonstrasikan bahwa penurunan temperature air, pH netral dan rendahnya salinitas, dalam kisaran yang biasanya terjadi secara alami, adalah faktor-faktor yang penting dalam meningkatkan infektivitas [25, 112, 150, 209]. Fluktuasi dalam temperatur [128], siklus beku-cair [212], dan konsentrasi ammonia [113] juga mengurangi durasi infektivitas. Faktor-faktor biologi tertentu pada habitat akuatik telah diperlihatkan memiliki pengaruh pada durasi infektivitas virus AI dalam air, mencakup filtrasi dan inaktivasi oleh kerrang [49] dan air aktif yang penuh secara biologic [42], dan kemungkinan bioakumulasi pada berudu [99] dan kutu air [1]. Bulu yang berkontak dengan feses atau air yang terkontaminasi juga dapat berlaku sebagai fomites tidak langsung untuk virus-virus ini [129]. Penelitian-penelitian ini memberi petunjuk adanya kompleksitas alasan mengapa virus AI bertahan lama di lingkungan, dimana pemahaman kita tentang hal ini masih belum lengkap.
Siklus maintenans virus Avian Influenza
Tidak diketahui bagaimana IAV dan keragaman subtipenya yang terdapat dalam sebuah populasi dipertahankan. Meskipun karakterisasi mekanisme-mekanisme ini merupakan tugas yang sangat kompleks dan menantang, sangat penting untuk memahami evolusi virus dan resiko yang terkait dengan kemunculan virus-virus penting seperti virus HPAI H5N1 Eurasia. Sangat mungkin bahwa siklus maintentas tergantung pada (i) kombinasi kontribusi transmisi dari burung ke burung melalui multi-hospes dan populasi unggas yang terpisah-pisah secara spasio-temporal, (ii) perilaku migrasi yang memberikan kesempatan hospes-hospes untuk bercampur, dan (iii) persistensi lingkungan pada habitat akuatik yang memungkinkan virus AI untuk terus bertahan dalam jangka pendek dan mungkin juga jangka panjang saat tidak terdapat hospes yang cocok. Siklus yang diusulkan ini didasarkan pada kriteria burung yang belum pernah terinfeksi sebelumnya/ rentan yang bisa jadi terinfeksi sepanjang tahun, dengan keterhubungan yang disediakan oleh adanya migrasi dan habitat yang dipergunakan bersama. Transmisi pada wilayah-wilayah breeding (berbiak), staging (tempat berhenti sementara dalam perjalanan saat migrasi), dan wintering (habitat/ tempat burung berada untuk menghindari musim dingin) telah ditentukan, begitu juga pergerakan virus AI ke arah selatan selama migrasi musim gugur dan ke arah utara selama migrasi musim semi. Bagian yang memiliki potensi menarik namun belum dimengerti dengan baik pada siklus ini berhubungan dengan kontribusi spesies individualnya, yang dapat berhubungan dengan tingkah laku migrasi ataupun tingkah laku spesifik lainnya. Hal ini telah tergambar pada burung blue-winged teal (Anas discors), yang merupakan migran awal yang tidak berada di wilayah utara saat prevalensi virus AI mencapai puncak pada spesies itik lainnya [207], dan untuk persamaan ekologisnya, itik Garganey (Anas querquedula) di Eropa dan Arika [40]. Tingkah laku migrasi ini dapat menyediakan populasi rentan untuk maintenans virus pada wilayah-wilayah wintering. Hubungan multispesies yang sama seperti ini juga bisa saja ada pada burung camar dan burung-burung pantai, dan bisa saja sebagiannya menjadi penghubung reservoir-reservoir ini dengan reservoir itik.
Sedangkan untuk reservoir lingkungan, isolasi banyak sekali subtipe virus AI dari air dan sedimen alami [187] dan demonstrasi persistensi virus dalam jangka yang sangat panjang di dalam air dibawah kondisi eksperimental memberikan dasar untuk hipotesis ini. Meskipun virus-virus ini mungkin bertahan pada habitat pembiakan unggas air dari tahun ke tahun, dan persistensi semacam itu diperlukan untuk maintenans virus yang sukses [18, 180], lebih mungkin bahwa persistensi lingkungan berlaku sebagai sumber jangka pendek bagi sisa-sisa virus yang menghubungkan kepergian dan kedatangan burung selama migrasi. Peranan potensial dalam mempertahankan virus di lingkungan membutuhkan penelitian yang lebih jauh, dan bisa saja penting dalam mendefinisikan bagaimana virus bergerak antar spesies burung migrasi dan spesies burung yang berpindah tempat secara local demikian juga populasi yang mungkin saja mempunyai kontak langsung secara terbatas atau terpisah secara temporal [86]. Hal ini juga mungkin sangat penting dalam menjelaskan bagaimana virus-virus ini bergerak diantara hospes liar dan domestik.
Transmisi LPAIV dari burung liar kepada unggas domestic sudah terdokumentasikan dengan baik, dan proses pertukaran virus LPAIV H5 dan H7 membuka jalan evolusi HPAIV [147]. Sebelum tahun 2002, hanya da satu laporan isolasi virus HPAI dari burung liar yang hidup bebas yang tidak diketahui berhubungan dengan unggas domestic yang terinfeksi. Ini termuat dalam laporan tahun 1961 yang melaporkan burung common tern yang terinfeksi H5N3 di Afrika Selatan [14]. Situasi ini berubah secara dramatis sebagai akibat wabah virus HPAI H5N1 di Eurasia. Pada tahun 2002 dan 2003, virus HPAI H5N1 telah diisolasi dari burung liar yang hidup bebas maupun dalam penangkaran di Hong Kong [48]. Selama tahun 2005 dan setelahnya, saat virus-virus ini tampaknya menyebar melalui migrasi bruung, isolasi irus HPAI H5N1 telah dilaporkan di Asia, Afrika dan Eropa [231]. Di Eropa, virus HPAI H5N1 telah dideteksi pada burung liar sampai tahun 2009 [50]; dan di Asia laporan terus berlanjut.
Meskipun telah banyak dugaan dilakukan, tidak ada bukti bahwa tumpahan (spillover) virus HPAI H5N1 burung liar akhirnya menjadikan burung liar sebagai reservoir untuk untuk virus-virus ini. Akan tetapi sekali virus ini masuk ke dalam populasi ini, maka banyak bukti yang menunjukkan bahwa introduksi virus HPAI berhubungan dengan migrasi burung liar dan pergerakannya secara lokal [188].
Spesies burung liar dimana virus HPAI H5N1 telah diisolasi meliputi spesies yang termasuk dalam Anseriformes, Charadriiformes, Ciconiiformes (herons (bangau) dan storks (bangau laut)), Columbiformes (burung merpati), Falconifromes (raptor), Galliformes (burung puyuh dan burung pegar), Gruiformes (coot dan moorhen), Passeriformes (burung yang bertengger di dahan/perching birds), Pelecaniformes (kormoran) dan Podicipediformes (grebes) [31, 48, 132, 137, 144, 188, 211, 231]. Spesies-spesies positif ini dapat digolongkan kedalam tiga kelompok secara umum. Kelompok pertama meliputi burung akuatik, dan kelompok ini utamanya disusun oleh burung dalam ordo Anseriformes. Kelompok kedua meliputi burung pemangsa, dan spesies lain seperti burung gagak yang dapat memangsa ataupun memakan bangkai burung lain (baik liar ataupun domestic). Kelompok terakhir meliputi banyak spesies burung pegar yang ada dalam lahan peridomestik / setengah liar setengah domestic. Terlibatnya burung air, terutama itik dan burung camar, konsisten dengan sejarah alami virus AI. Akan tetapi, spesies yang terlibat meliputi angsa dan itik penyelam dalam jumlah yang jauh lebih kecil. Kelompok burung air ini kurang terepresentasikan dalam hubungannya dengan isolasi virus LPAI yang dilaporkan. Spesies burung liar pemangsa dan pemakan bangkau umumnya tidak dihubungkan dengan virus LPAI yang terjadi secara alami, dan hal ini mungkin berhubungan dengan kurangnya morbiditas dan mortalitas pada burung-burung yang terinfeksi virus LPAI yang berperan sebagai mangsa dan makanan potensial bagi spesies-speseis tersebut. Pada Burung pegar, yang biasanya tidak dihubungkan dengan infeksi LPAI, sebagian besar infeksi mungkin merupakan ahsil dari burung hasil penangkaran yang dijual di pasar burung, atau langsung terekspos dengan flok unggas yang terinfeksi [200].
Telah banyak percobaan infeksi virus HPAI H5N1 kepada spesies burung liar [19, 21-23, 71-73, 111, 131, 152, 163, 167-169, 216], dan studi-studi ini secara konsisten juga telah memperlihatkan kisaran tingkat kerentanan dan viral shedding yang luas yang utamanya dihubungkan dengan infeksi sistem respirasi dibandingkan dengan infeksi saluran pencernaan. Respons klinis pada itik dan burung camar berhubungan dengan garis keturunan virus HPAI H5N1 [19, 168, 216], dan hasl akhir klinisnya dapat bervariasi diantara spesies-spesies yang berhubungan dekat secara taksonomi. Hal ini diperlihatkan baik pada spesies-spesies itik Amerika Utara, meliputi entok (mallard), blue-winged teal, redhead (Aythya Americana), wood duck (Aix sponsa), dan Northers pintail (Anas acuta) [19], dan pada spesies-spesies itik Eurasia, meliputi tufted duck (Aythya fuligula), common pochard (Aythya ferina), entok, common teal, Eurasian widgeon (Anas Penelope), dan gadwaal (Anas Strepera) [111], juga pada angsa besar dan angsa [22]. Variasi taraf dan durasi viral shedding juga sangat bervariasi antar spesies, akan tetapi titer virus (terutama dari swab kloaka) umumnya lebih rendah jika dibandingkan dengan virus LPAI. Titer virus terobservasi berhubungan dengan respons klinism dengan titer tertinggi dihubungkan dengan burung yang mengalami keparahan gejala klinis paling parah [19, 111, 216]. Pada angsa, variasi antar spesies berhubungan dengan waktu onset penyakit klinis dan durasi dan taraf viral shedding juga tampak jelas [22]. Terdapat beberapa penelitian terbatas tebtabf enfek pre-paparane terhadap infeksi HPAI H5N1 [15, 36], dan hasilnya menunjukkan bahwa pre-paparan dengan virus LPAI homolog dan hetrolog dapat mempengaruhi baik tingkat keparahan klinis dan viral shedding.
Untuk virus LPAI, resiko infeksi dan potensi maintenance virus pada reservoir unggas tergantung pada tingkat kerentanan dan potensi kontak, dan yang terakhir ini dapat sangat dipengaruhi oleh tingkah laku spesies. Dengan virus-virus baru seperti virus HPAI H5N1, resiko pergerakan virus pada individu dan populasi perlu untuk dimengerti, dan untuk kasus ini, kisaran hospes, ukuran populasi, dan tingkah laku migrasi menjadi sangat penting [50, 63, 175]. Banyak penelitian yang berhubungan virus HPAI H5N1 baik berdasarkan ornitologi atau telah menggunakan data dan teknik ornitologi secara ekstensif. Penggunaan telemetri satelit adalah sebuah contoh bagus. Banyak studi telah menggunakan telemetri [66], terkadang besamaan dengan pengujian influenza, untuk menentukan poin kontak potensial dengan flok unggas yang terinfeksi [218], untuk menghubungkan atau tidak menghubungkan area dimana wabah telah terjadi pada unggas [60, 171], dan untuk menentukan kemampuan spesies hospes spesifik dalam memindahkan virus-virus ini [154].
Secara kolektif, seluruh studi virus HPAI H5N1 secara eksperimental maupun lapangan jelas mengindikasikan bahwa virus-virus ini dapat bergerak melalui populasi burung migrasi, dan berkontribusi terhadap penyebaran virus secara local [65], dan bahwa burung peridomestik dapat terinfeksi dalam skala local. Studi-studi ini tidak menyokong penetapan reservoir hewan liar untuk virus-virus ini, dan lebih mungkin bahwa itik domestik merepresentasikan reservoir sejati virus ini daripada itik liar [30, 103]. Kegagalan virus-virus ini untuk menetap pada reservoir burung liar sangat mungkin berhubungan dengan adaptasinya kepada hospes domestic dan lingkungan mereka, pemilihan waktu migrasi [125], dan mungkin dilusi atau imunitas populasi, karena populasi liar ini sudah merupakan rumah bagi banyak virus LPAI. Setelah semua itu, mungkin bahwa keterlibatan burung liar dalam epidemiologi virus-virus ini dapat diabaikan jika tidak ada kontak dengan unggas domestic yang terinfeksi.
Virus ini bukanlah virus HPAI pertama yang mampu bereplikasi pada spesies burung liar, dan dengan virus yang lebih baru lagi seperti virus HPAI H5N8, yang telah diisolasi dari burung liar dan unggas domestik di Asia [133], Eropa dan Amerika, dan virus LPAI H7N9 yang telah menginfeksi manusia di China [239], pertanyaan-pertanyan terkait dengan terlibatnya burung liar berkenaan dengan virus tersebut akan terus muncul.
Pada saat ini pemahaman kita tentang AIV pada reservoir unggas alami sangat ekstensif, akan tetapi belum lagi mendekati lengkap. Seperti yang diperlihatkan pada Gambar 7.1, terdapat banyak variable yang berpotensi mempengaruhi dan mungkin juga meregulasi virus-virus ini diantara populasi virus AI, hospes unggas, dan lingkungan. Pemahaman hubungan interaksi ini diperlukan untuk menjawab pertanyaan-pertanyaan dasar yang berhubungan dengan mempertahankan virus pada populasi burung liar dalam hubungannya dengan system alami maupun system buatan manusia, untuk mengerti keberagaman subtipe, untuk menilai potensi-potensinya dalam menentukan keberhasilan invasi virus AI ke hospes baru ataupun wilayah-wilayah geografis baru, dan untuk mengerti resiko-resiko yang berhubungan dengan pergerakan virus-virus ini kepada hospes hewan domestic ataupun manusia.
Diantara faktor hospes yang telah disebutkan, imunitas populasi adalah yang paling sedikit dimengerti dan membutuhkan penelitian lebih lanjut. Efek-efek potensial yang berhubungan dengan struktur komunitas, struktur populasi, dan migrasi juga penting, dan penelitian yang telah ada saat ini belum memadai. Efek-efek ini bersifat penting dan kritikal dalam memahami epidemiologi virus-virus ini yang dapat memiliki kesamaan baik pada populasi burung liar maupun unggas domestic.
Populasi virus AI telah didefinisikan dengan baik, dan signifikansi subtipe dimengerti dalam hubungannya dengan pathogenesis (H5/H7), batasan hospes (H13/H16), atau predominansinya (H3/H4). Meskipun begitu, belum diketahui mengapa subtipe-subtipe ini berevolusi dan bagaimana mereka bertahan di dalam tubuh satu individu. Barug-burung liar, teutama itik dan burung camar, memberikan kesempatan unik untuk mengerti tentang betapa beragamnya interaksi virus AI dan kompetisinya dalam suatu populasi. Dalam perspektif penerapan, pemahaman tentang faktor-faktor lain seperti subtipe musiman dan assosiasi hospes dapat lebih meningkatkan efisiensi upaya surveilans di masa depan. Keragaman genetic yang ada pada ppol gen virus AI pada burung liar juga telah didokumentasikan dengan baik, dan ada banyak penelitian yang mendemonstrasikan pergerakan gen virus antar benua serta kesuksesan masuknya gen itu. Ana tetapi, karena faktor-faktor yang terkait dengan dengan kesuksesan pemasukan virus ini belum diketahui, kita belum punya pemahaman yang lengkap tentang resiko-resiko terkait dengan pergerakan virus-virus ini secara spesifik seperti pada H5N1 Eurasia.
Peranan persisnya dari persistensi lingkungan dan reservoir lingkungan potensial masih bersifat spekulatif, akan tetapi tetaplah penting. Jika persistensinya bersifat jangka panjang, ‘tingkah laku’ ini dapat mempengaruhi baik struktur genetic dari populasi virus AI maupun distribusi spasial dan temporal virus-virus ini pada populasi hospes. Jika persistensinya di lingkungan pada waktu yang pendek, mungkin saja itu merupakan komponen penting dalam siklus maintenans virus baik HPAI maupun HPAI. Relatif sedikit penelitian yang mengkaji tentang potensi faktor abiotic dan biotik yang mempengaruhi persistensi lingkungan vius-virus ini. sebagai tambahan lagi, penelitian berdasarkan uji lapangan atau system lingkungan yang lebih kompleks yang lebih dapat merefleksikan system alami sangat diperlukan dalam rangka untuk memahami subyek ini dengan lebih baik.
Akhirnya, karena cepatnya perubahan lingkungan dan populasi yang dihubungkan dengan burung-burung liar dan domestic di seluruh dunia, pengetahuan kita tentang epidemiologi virus AI akan terus berevolusi dan menantang pemahaman kita tetang system dan interaksinya yang kompleks. Hal ini terutama benar pada wilayah dimana terdapat meningkatnya kontak antara populasi liar dan domestik.
Abbas, M. D., J. Nazir, P. Stumpf and R. E. Marschang, 2012: Role of Water Fleas (Daphniamagna) in the Accumulation of Avian Influenza Viruses from the Surrounding Water. Intervirology,55, 365-371.
Abenes, G. B., K. Okazaki, H. Fukushi, H. Kida, E. Honda, K. Yagyu, M. Tsuji, H. Sato, E. Ono and R. Yanagawa, 1982: Isolation of ortho-and paramyxoviruses from feral birds in Hokkaido, Japan–1980-1981. Japanese Journal of Veterinary Science,44, 703-708.
Alexander, D. J., W. W. Allan and J. W. Harkness, 1974: Isolation of influenza virus from psittacines. Research in Veterinary Science,17, 125-127.
Alexander, D., G. Parsons and R. Manvell, 1986: Experimental assessment of the pathogenicity of eight avian influenza A viruses of H5 subtype for chickens, turkeys, ducks and quail. Avian Pathology,15, 647-662.
Alexander, D., W. Allan, D. Parsons and G. Parsons, 1978: The pathogenicity of four avian influenza viruses for fowls, turkeys and ducks. Research in Veterinary Science,24, 242-247.
Alfonso, C. P., B. S. Cowen and H. V. Campen, 1995: Influenza A viruses isolated from waterfowl in two wildlife management areas of Pennsylvania. Journal of Wildlife Diseases,31, 179-185.
Amin, A., M. Shalaby and I. Z. Imam, 1980: Studies on influenza virus isolated from migrating birds in Egypt. Comparative Immunology, Microbiology Infectious Diseases,3, 241-246.
Arnal, A., M. Vittecoq, J. Pearce-Duvet, M. Gauthier-Clerc, T. Boulinier and E. Jourdain, 2015: Laridae: A neglected reservoir that could play a major role in avian influenza virus epidemiological dynamics. Critical Reviews in Microbiology,41, 508-519.
Asplin, F. D., 1970: Examination of sera from wildfowl for antibodies against the viruses of duck plague, duck hepatitis and duck influenza. The Veterinary Record,87, 182-183.
Bahl, A. K., B. S. Pomeroy, B. C. Easterday and S. Mangundimedjo, 1975: Isolation of type A influenza viruses from the migratory waterfowl along the Mississippi flyway. Journal of Wildlife Diseases,11, 360-363.
Bahl, A. K., B. S. Pomeroy, S. Mangundimedjo and B. C. Easterday, 1977: Isolation of type A influenza and Newcastle disease viruses from migratory waterfowl in the Mississippi flyway. Journal of the American Veterinary Medical Association,171, 949-951.
Bahl, J., D. Vijaykrishna, E. C. Holmes, G. J. Smith and Y. Guan, 2009: Gene flow and competitive exclusion of avian influenza A virus in natural reservoir hosts. Virology,390, 289-297.
Barton, H. D., P. Rohani, D. E. Stallknecht, J. Brown and J. M. Drake, 2014: Subtype diversity and reassortment potential for co‐circulating avian influenza viruses at a diversity hot spot. Journal of Animal Ecology,83, 566-575.
Becker, W., 1966: The isolation and classification of tern virus: influenza virus A/tern/South Africa/1961. Journal of Hygine,64, 309-320.
Berhane, Y., C. Embury-Hyatt, M. Leith, H. Kehler, M. Suderman and J. Pasick, 2014: Pre-exposing Canada Geese (Branta canadensis) to a low-pathogenic H1N1 avian influenza virus protects them against H5N1 HPAI virus challenge. Journal of wildlife diseases,50, 84-97.
Boudreault, A., J. Lecomte and V. Hinshaw, 1980: Antigenic characterization of influenza A virus isolated from birds captured in Ontario, Quebec, and the maritime provinces during the 1977 season. Revue canadienne de biologie,39, 107-114.
Boyce, W. M., S. Schobel, V. G. Dugan, R. Halpin, X. Lin, D. E. Wentworth, L. L. Lindsay, E. Mertens and M. Plancarte, 2013: Complete genome sequence of a reassortant H14N2 avian influenza virus from California. Genome Announcements,1, e00543-00513.
Breban, R., J. M. Drake, D. E. Stallknecht and P. J. P. c. b. Rohani, 2009: The role of environmental transmission in recurrent avian influenza epidemics. PLoS Computational Biology,5,
Brown, J. D., D. E. Stallknecht, J. R. Beck, D. L. Suarez and D. E. Swayne, 2006: Susceptibility of North American ducks and gulls to H5N1 highly pathogenic avian influenza viruses. Emerging Infectious Diseases,12,
Brown, J. D., D. E. Swayne, R. J. Cooper, R. E. Burnes and D. E. Stallknecht, 2007: Persistence of H5 and H7 influenza viruses in water. Avian Diseases,51, 285-289.
Brown, J. D., D. E. Stallknecht, S. Valeika and D. E. Swayne, 2007: Susceptibility of wood ducks to highly pathogenic avian influenza viruses. Journal of Wildlife Diseases,43.
Brown, J. D., D. E. Stallknecht and D. E. Swayne, 2008: Experimental infection of swans and geese with highly pathogenic avian influenza virus (H5N1) of Asian lineage. Emerging Infectious Diseases,14,
Brown, J. D., D. E. Stallknecht, R. D. Berghaus and D. E. Swayne, 2009: Infectious and lethal doses of H5N1 highly pathogenic avian influenza virus for house sparrows (Passer domesticus) and rock pigeons (Columbia livia). Journal of Veterinary Diagnostic Investigation,21, 437-445.
Brown, J. D., D. E. Stallknecht, R. D. Berghaus and D. E. Swayne, 2009: Evaluation of a commercial blocking enzyme-linked immunosorbent assay to detect avian influenza virus antibodies in multiple experimentally infected avian species. Clinical and Vaccine Immunology,16.
Brown, J. D., G. Goekjian, R. Poulson, S. Valeika and D. E. Stallknecht, 2009: Avian Influenza in water; infectivity is dependent on pH, salinity, and temperature. Veterinary Microbiology,136:20-26.
Brown, J., R. Poulson, D. Carter, C. Lebarbenchon, M. Pantin-Jackwood, E. Spackman, E. Shepherd, M. Killian and D. Stallknecht, 2012: Susceptibility of avian species to North American H13 low pathogenic avian influenza viruses. Avian Diseases,56, 969-975.
Brown, J. D., R. D. Berghaus, T. P. Costa, R. Poulson, D. L. Carter, C. Lebarbenchon and D. E. Stallknecht, 2012: Intestinal excretion of a wild bird-origin H3N8 low pathogenic avian influenza virus in mallards (Anas platyrhynchos). Journal of Wildlife Diseases,48, 991-998.
Busquets, N., A. Alba, S. Napp, A. Sánchez, E. Serrano, R. Rivas, J. I. Núñez and N. Majó, 2010: Influenza A virus subtypes in wild birds in North-Eastern Spain (Catalonia). Virus Research,149, 10-18.
Campbell, D., 1999: Gull mortality—Kitchener. Wildlife Health Centre Newsletter,6, 8-9.
Capelle, J., D. Zhao, M. Gilbert, I. Nelson, S. H. Newman, J. Y. Takekawa, N. Gaidet, D. J. Prosser, Y. Liu, P. Li, Y. Shu and X. Xiao, 2014: Risks of avian influenza transmission in areas of intensive free-ranging duck production with wild waterfowl. EcoHealth,11, 109-119.
Chen, H., G. Smith, S. Zhang, K. Qin, J. Wang, K. Li, R. Webster, J. Peiris and Y. Guan, 2005: H5N1 virus outbreak in migratory waterfowl. Nature,436, 191-192.
Cheng, M., M. Lee, Y. Ho, W. Chyi and C. Wang, 2010: Avian influenza monitoring in migrating birds in Taiwan during 1998–2007. Avian Diseases,54, 109-114.
Costa, T. P., J. D. Brown, E. W. Howerth and D. E. Stallknecht, 2010: The effect of age on avian influenza viral shedding in mallards (Anas platyrhynchos). Avian Diseases,54, 581-585.
Costa, T. P., J. D. Brown, E. W. Howerth and D. E. Stallknecht, 2010: Effect of a prior exposure to a low pathogenic avian influenza virus in the outcome of a heterosubtypic low pathogenic avian influenza infection in mallards (Anas platyrhynchos). Avian Diseases,54, 1286-1291.
Costa, T. P., J. D. Brown, E. W. Howerth and D. E. Stallknecht, 2011: Variation in viral shedding patterns between different wild bird species infected experimentally with low-pathogenicity avian influenza viruses that originated from wild birds. Avian Pathology,40, 119-124.
Costa, T. P., J. D. Brown, E. W. Howerth, D. E. Stallknecht and D. E. J. P. O. Swayne, 2011: Homo-and heterosubtypic low pathogenic avian influenza exposure on H5N1 highly pathogenic avian influenza virus infection in wood ducks (Aix sponsa). 6,
Cumming, G. S., A. Caron, C. Abolnik, G. Cattoli, L. W. Bruinzeel, C. E. Burger, K. Cecchettin, N. Chiweshe, B. Mochotlhoane and G. L. Mutumi, 2011: The ecology of influenza A viruses in wild birds in southern Africa. EcoHealth,8, 4-13.
Curran, J. M., T. M. Ellis and I. D. Robertson, 2014: Surveillance of Charadriiformes in northern Australia shows species variations in exposure to avian influenza virus and suggests negligible virus prevalence. Avian Diseases,58, 199-204.
De Marco, M., G. Foni, L. Campitelli, E. Raffini, L. Di Trani, M. Delogu, V. Guberti, G. Barigazzi and I. Donatelli, 2003: Circulation of influenza viruses in wild waterfowl wintering in Italy during the 1993–99 period: evidence of virus shedding and seroconversion in wild ducks. Avian Diseases,47, 861-866.
De Marco, M. A., E. Foni, I. Campitelli, M. Delogu, E. Raffini, C. Chiapponi, G. Barigazi, P. Cordioli, L. Di Trani and I. Donnatelli, 2005: Influenza virus circulation in wild aquatic birds in Italy during H5N2 and H7N1 poultry epidemic periods (1998 to 2000). Avian Pathology,34, 480-485.
Deibel, R., D. E. Emord, W. Dukelow, V. S. Hinshaw and J. M. Wood, 1985: Influenza viruses and paramyxoviruses in ducks in the Atlantic flyway, 1977-1983, including an H5N2 isolate related to the virulent chicken virus. Avian Diseases, 970-985.
Domanska-Blicharz, K., Z. Minta, K. Smietanka, S. Marché and T. Van Den Berg, 2010: H5N1 high pathogenicity avian influenza virus survival in different types of water. Avian Diseases,54, 734-737.
Donis, R. O., W. J. Bean, Y. Kawaoka and R. G. Webster, 1989: Distinct lineages of influenza virus H4 hemagglutinin genes in different regions of the world. Virology,169, 408-417.
Downie, J. C. and W. G. Laver, 1973: Isolation of a type A influenza virus from an Australian pelagic bird. Virology,51, 259-269.
Downie, J. C., V. Hinshaw and W. Laver, 1977: The ecology of influenza: isolation of type ‘A’influenza viruses from Australian pelagic birds. Australian Journal of Experimental Biology Medical Science,55, 635-643.
Dugan, V. G., R. Chen, D. J. Spiro, N. Sengamalay, J. Zaborsky, E. Ghedin, J. Nolting, D. E. Swayne, J. A. Runstadler and G. M. Happ, 2008: The evolutionary genetics and emergence of avian influenza viruses in wild birds. PLoS Pathogens,4,
Dusek, R. J., G. T. Hallgrimsson, H. S. Ip, J. E. Jónsson, S. Sreevatsan, S. W. Nashold, J. L. TeSlaa, S. Enomoto, R. A. Halpin and X. Lin, 2014: North Atlantic migratory bird flyways provide routes for intercontinental movement of avian influenza viruses. PLoS One,9,
Ellis, T. M., R. Barry Bousfield, L. A. Bissett, K. C. Dyrting, G. S. Luk, S. Tsim, K. Sturm-Ramirez, R. G. Webster, Y. Guan and J. M. J. A. P. Peiris, 2004: Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. 33, 492-505.
Faust, C., D. Stallknecht, D. Swayne and J. Brown, 2009: Filter-feeding bivalves can remove avian influenza viruses from water and reduce infectivity. Proceedings of the Royal Society B: Biological Sciences,276, 3727-3735.
Feare, C. J., 2010: Role of wild birds in the spread of highly pathogenic avian influenza virus H5N1 and implications for global surveillance. Avian Diseases,54, 201-212.
Fereidouni, S. R., E. Starick, M. Beer, H. Wilking, D. Kalthoff, C. Grund, R. Häuslaigner, A. Breithaupt, E. Lange and T. C. Harder, 2009: Highly pathogenic avian influenza virus infection of mallards with homo-and heterosubtypic immunity induced by low pathogenic avian influenza viruses. PLoS One,4,
Ferro, P. J., C. M. Budke, M. J. Peterson, D. Cox, E. Roltsch, T. Merendino, M. Nelson and B. Lupiani, 2010: Multiyear surveillance for avian influenza virus in waterfowl from wintering grounds, Texas coast, USA. Emerging Infectious Diseases,16,
Fouchier, R. A. M., B. Olsen, T. M. Bestebroer, S. Herfst, L. Van der Kemp, G. F. Rimmelzwaan and A. D. M. E. Osterhaus, 2003: Influenza A virus surveillance in wild birds in Northern Europe in 1999 and 2000. Avian Diseases,47, 857-860.
Fouchier, R. A. M., V. Munster, A. Wallensten, T. M. Bestebroer, S. Herfst, D. Smith, G. F. Rimmelzwaan, B. r. Olsen and A. D. M. E. Osterhaus, 2005: Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. Journal of Virology,79, 2814-2822.
Franca, M., D. E. Stallknecht and E. W. Howerth, 2013: Expression and distribution of sialic acid influenza virus receptors in wild birds. Avian Pathology,42, 60-71.
França, M., R. Poulson, J. Brown, E. W. Howerth, R. D. Berghaus, D. Carter and D. E. Stallknecht, 2012: Effect of different routes of inoculation on infectivity and viral shedding of LPAI viruses in mallards. Avian Diseases,56, 981-985.
Fries, A. C., J. M. Nolting, A. Danner, R. G. Webster, A. S. Bowman, S. Krauss and R. D. Slemons, 2013: Evidence for the circulation and inter-hemispheric movement of the H14 subtype influenza A virus. PLoS One,8,
Fusaro, A., I. Monne, G. Cattoli, R. De Nardi, A. Salviato, A. M. Martin, I. Capua and C. Terregino, 2010: Gene segment reassortment between Eurasian and American clades of avian influenza virus in Italy. Archives of Virology,155, 77-81.
Gaidet, N., T. Dodman, A. Caron, G. Balança, S. Desvaux, F. Goutard, G. Cattoli, F. Lamarque, W. Hagemeijer and F. Monicat, 2007: Avian influenza viruses in water birds, Africa. Emerging Infectious Diseases,13,
Gaidet, N., S. H. Newman, W. Hagemeijer, T. Dodman, J. Cappelle, S. Hammoumi, L. De Simone and J. Y. Takekawa, 2008: Duck migration and past influenza A (H5N1) outbreak areas. Emerging Infectious Diseases,14,
Gaidet, N., A. B. Ould El Mamy, J. Cappelle, A. Caron, G. S. Cumming, V. Grosbois, P. Gil, S. Hammoumi, R. S. De Almeida and S. R. Fereidouni, 2012: Investigating avian influenza infection hotspots in old-world shorebirds. PLoS One,7,
Gaidet, N., A. Caron, J. Cappelle, G. S. Cumming, G. Balança, S. Hammoumi, G. Cattoli, C. Abolnik, R. Servan de Almeida and P. Gil, 2012: Understanding the ecological drivers of avian influenza virus infection in wildfowl: a continental-scale study across Africa. Proceedings of the Royal Society B: Biological Sciences,279, 1131-1141.
Gale, P., A. Goddard, A. C. Breed, R. M. Irvine, L. Kelly and E. L. Snary, 2014: Entry of H 5 N 1 highly pathogenic avian influenza virus into E urope through migratory wild birds: a qualitative release assessment at the species level. Journal of Applied Microbiology,116, 1405-1417.
Garamszegi, L. Z. and A. P. Møller, 2007: Prevalence of avian influenza and host ecology. Proceedings of the Royal Society B: Biological Sciences,274, 2003-2012.
Gauthier‐Clerc, M., C. Lebarbenchon and F. Thomas, 2007: Recent expansion of highly pathogenic avian influenza H5N1: a critical review. Ibis,149, 202-214.
Gilbert, M., S. H. Newman, J. Y. Takekawa, L. Loth, C. Biradar, D. J. Prosser, S. Balachandran, M. V. S. Rao, T. Mundkur and B. Yan, 2010: Flying over an infected landscape: distribution of highly pathogenic avian influenza H5N1 risk in South Asia and satellite tracking of wild waterfowl. EcoHealth,7, 448-458.
Gonzalez-Reiche, A. S. and D. R. Perez, 2012: Where do avian influenza viruses meet in the Americas? Avian Diseases,56, 1025-1033.
Gorman, O. T., W. J. Bean and R. G. Webster, 1992: Evolutionary processes in influenza viruses: divergence, rapid evolution, and stasis. Current Topics in Microbilogy and Immunology,176, 75-97.
Graves, I. L., 1992: Influenza viruses in birds of the Atlantic flyway. Avian Diseases, 1-10.
Gresikova, M., B. Tůmová, A. Stumpa and M. Sekeyova, 1978: Isolation of influenza virus from wild ducks (Anas platyrhynchos). Acta Virologica,22, 296-301.
Hall, J. S., H. S. Ip, J. C. Franson, C. Meteyer, S. Nashold, J. L. TeSlaa, J. French, P. Redig and C. Brand, 2009: Experimental infection of a North American raptor, American kestrel (Falco sparverius), with highly pathogenic avian influenza virus (H5N1). PLoS One,4,
Hall, J. S., J. C. Franson, R. E. Gill, C. U. Meteyer, J. L. TeSlaa, S. Nashold, R. J. Dusek and H. S. Ip, 2011: Experimental challenge and pathology of highly pathogenic avian influenza virus H5N1 in dunlin (Calidris alpina), an intercontinental migrant shorebird species. Influenza and Other Respiratory Viruses,5, 365-372.
Hall, J. S., J. L. TeSlaa, S. W. Nashold, R. A. Halpin, T. Stockwell, D. E. Wentworth, V. Dugan and H. S. Ip, 2013: Evolution of a reassortant North American gull influenza virus lineage: drift, shift and stability. Virology Journal,10, 1-9.
Halvorson, D., D. Karunakaran, D. Senne, C. Kelleher, C. Bailey, A. Abraham, V. Hinshaw and J. Newman, 1983: Simultaneous monitoring of sentinel ducks and turkeys in Minnesota. Avian Diseases,27, 77-85.
Halvorson, D., D. Karunakaran, D. Senne, C. Kelleher, C. Bailey, A. Abraham, V. Hinshaw and J. Newman, 1985: Epizootiology of avian influenza: simultaneous monitoring of sentinel ducks and turkeys in Minnesota. Applied and Enviromental Microbiology,49, 914-919.
Hannoun, C., 1977: Isolation from birds of influenza viruses with human neuraminidase. Developments in Biological Standardization,39, 469-472.
Hannoun, C. and J. M. Devaux, 1980: Circulation of influenza viruses in the bay of the Somme river (author’s transl). Comparative Immunology, Microbiology and Infectious Diseases,3, 177-183.
Hanson, B. A., D. E. Stallknecht, D. E. Swayne, L. A. Lewis and D. A. Senne, 2003: Avian influenza viruses in Minnesota ducks during 1998–2000. Avian Diseases,47 (Suppl.), 867-871.
Hanson, B. A., M. P. Luttrell, V. H. Goekjian, L. Niles, D. E. Swayne, D. A. Senne and D. E. Stallknecht, 2008: Is the occurrence of avian influenza virus in Charadriiformes species and location dependent? Journal of Wildlife Diseases,44, 351-361.
Hanson, B. A., D. E. Swayne, D. A. Senne, D. S. Lobpries, J. Hurst and D. E. Stallknecht, 2005: Avian influenza viruses and paramyxoviruses in wintering and resident ducks in Texas. Journal of Wildlife Diseases,41, 624-628.
Harris, M. T., J. D. Brown, V. H. Goekjian, M. P. Luttrell, R. L. Poulson, B. R. Wilcox, D. E. Swayne and D. E. Stallknecht, 2010: Canada geese and the epidemiology of avian influenza viruses. Journal of Wildlife Diseases,46, 981-987.
Hatchette, T. F., D. Walker, C. Johnson, A. Baker, S. P. Pryor and R. G. Webster, 2004: Influenza A viruses in feral Canadian ducks: extensive reassortment in nature. Journal of General Virology,85, 2327-2337.
Hénaux, V. and M. D. Samuel, 2011: Avian influenza shedding patterns in waterfowl: implications for surveillance, environmental transmission, and disease spread. Journal of Wildlife Diseases,47, 566-578.
Hill, N. J., J. Y. Takekawa, C. J. Cardona, J. T. Ackerman, A. K. Schultz, K. A. Spragens and W. M. Boyce, 2010: Waterfowl ecology and avian influenza in California: do host traits inform us about viral occurrence? Avian Diseases,54, 426-432.
Hill, N. J., J. Y. Takekawa, C. J. Cardona, B. W. Meixell, J. T. Ackerman, J. A. Runstadler and W. M. Boyce, 2012: Cross-seasonal patterns of avian influenza virus in breeding and wintering migratory birds: a flyway perspective. Vector-Borne Zoonotic Diseases,12, 2243-2253.
Hill, N. J., J. Y. Takekawa, J. T. Ackerman, K. A. Hobson, G. Herring, C. J. Cardona, J. A. Runstadler and W. M. Boyce, 2012: Migration strategy affects avian influenza dynamics in mallards (Anas platyrhynchos). Molecular Ecology,21, 5986-5999.
Hinshaw, V. S., R. G. Webster and B. Turner, 1978: Novel influenza A viruses isolated from Canadian feral ducks: including strains antigenically related to swine influenza (Hsw1N1) viruses. Journal of General Virology,41, 115-127.
Hinshaw, V. S., R. G. Webster and B. Turner, 1979: Water-borne transmission of influenza A viruses? Intervirology,11, 66-68.
Hinshaw, V. S., R. G. Webster and B. Turner, 1980: The perpetuation of orthomyxoviruses and paramyxoviruses in Canadian waterfowl. Canadian Journal of Microbiology,26, 622-629.
Hinshaw, V. S., G. M. Air, A. Gibbs, L. Graves, B. Prescott and D. Karunakaran, 1982: Antigenic and genetic characterization of a novel hemagglutinin subtype of influenza A viruses from gulls. Journal of Virology,42, 865-872.
Hinshaw, V. S. and R. G. Webster, 1982: The natural history of influenza A viruses. In: Basic Applied Influenza Research. CRC Press, Boca Raton, Florida, FL. 79-104.
Hinshaw, V. S., J. M. Wood, R. G. Webster, R. Deibel and B. Turner, 1985: Circulation of influenza viruses and paramyxoviruses in waterfowl originating from two different areas of North America. Bulletin of the World Health Organization,63,
Hinshaw, V. S., V. F. Nettles, L. F. Schorr, J. M. Wood and R. G. Webster, 1986: Influenza virus surveillance in waterfowl in Pennsylvania after the H5N2 avian outbreak. Avian Diseases, 207-212.
Hjulsager, C. K., S. Ø. Breum, R. Trebbien, K. J. Handberg, O. R. Therkildsen, J. J. Madsen, K. Thorup, J. A. Baroch, T. J. DeLiberto and L. E. Larsen, 2012: Surveillance for avian influenza viruses in wild birds in Denmark and Greenland, 2007–10. Avian Diseases,56, 992-998.
Hlinak, A., R. U. Mühle, O. Werner, A. Globig, E. Starick, H. Schirrmeier, B. Hoffmann, A. Engelhardt, D. Hübner and F. J. Conraths, 2006: A virological survey in migrating waders and other waterfowl in one of the most important resting sites of Germany. Journal of Veterinary Medicine. B. Infectious Diseases and Veterinary Public Health,53, 105-110.
Homme, P. J. and B. C. Easterday, 1970: Avian influenza virus infections. IV. Response of pheasants, ducks, and geese to Influenza A/Turkey/Wisconsin/1966 virus. Avian Diseases,14, 285-290.
Honda, E., H. Kida, R. Yanagawa, Y. Matsuura, K. Yagyu, M. Tsuji, K. Ueno, N. Yamauchi, S. Mishima and H. Ogi, 1981: Survey of influenza viruses in feral birds in 1979 and isolation of a strain possessing Hav6Nav5 from cloaca of an eastern dunlin. Japanese Journal of Veterinary Research,29, 83-87.
Horm, S. V., R. A. Gutiérrez, S. Sorn and P. Buchy, 2012: Environment: a potential source of animal and human infection with influenza A (H5N1) virus. Influenza and Other Respiratory Viruses,6, 442-448.
Hoye, B. J., V. J. Munster, H. Nishiura, M. Klaassen and R. A. M. Fouchier, 2010: Surveillance of wild birds for avian influenza virus. Emerging Infectious Diseases,16,
Hoye, B. J., V. J. Munster, H. Nishiura, R. A. Fouchier, J. Madsen and M. Klaassen, 2011: Reconstructing an annual cycle of interaction: natural infection and antibody dynamics to avian influenza along a migratory flyway. Oikos,120, 748-755.
Hua, Y. P., H. L. Chai, S. Y. Yang, X. W. Zeng and Y. Sun, 2005: Primary survey of avian influenza virus and Newcastle disease virus infection in wild birds in some areas of Heilongjiang Province, China. Journal of Veterinary Science,6, 311-315.
Hulse-Post, D. J., K. M. Sturm-Ramirez, J. Humberd, P. Seiler, E. A. Govorkova, S. Krauss, C. Scholtissek, P. Puthavathana, C. Buranathai and T. D. Nguyen, 2005: Role of domestic ducks in the propagation and biological evolution of highly pathogenic H5N1 influenza viruses in Asia. Proceedings of the National Academy of Sciences,102, 10682-10687.
Ibrahim, H. M., I. P. R. Awang, D. J. Alexander, R. J. Manvell, I. Aini and A. L. Ibrahim, 1990: Isolations of influenza A viruses from passerine birds in Malaysia. Veterinary Record,127.
Iftimovici, R. and P. Al, 1980: Isolation of influenza virus A/USSR 90/77 (H1N1) from wild birds. Virologie,31,
Irwin, C. K., K.-J. Yoon, C. Wang, S. J. Hoff, J. J. Zimmerman, T. Denagamage and A. M. O’Connor, 2011: Using the systematic review methodology to evaluate factors that influence the persistence of influenza virus in environmental matrices. Applied and Enviromental Microbiology,77, 1049-1060.
Isachenko, V. A., L. Y. Zakstelskaya, I. G. Roslaya, L. D. Odinok, E. V. Molibog and D. K. Lvov, 1974: Strains similar to Hong-Kong variant of influenza virus isolated from synanthropic and wild migrating birds. Ivanovsky Institute of Virology,2, 156-165.
Ito, T., K. Okazaki, Y. Kawaoka, A. Takada, R. Webster and H. Kida, 1995: Perpetuation of influenza A viruses in Alaskan waterfowl reservoirs. Archives of Virology,140, 1163-1172.
Jourdain, E., G. Gunnarsson, J. Wahlgren, N. Latorre-Margalef, C. Bröjer, S. Sahlin, L. Svensson, J. Waldenström, Å. Lundkvist and B. Olsen, 2010: Influenza virus in a natural host, the mallard: experimental infection data. PLoS One,5,
Kawaoka, Y., T. M. Chambers, W. L. Sladen and R. Gwebster, 1988: Is the gene pool of influenza viruses in shorebirds and gulls different from that in wild ducks? Virology,163, 247-250.
Keawcharoen, J., D. Van Riel, G. van Amerongen, T. Bestebroer, W. E. Beyer, R. Van Lavieren, A. D. M. E. Osterhaus, R. A. M. Fouchier and T. Kuiken, 2008: Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1). Emerging Infectious Diseases,14,
Keeler, S. P., R. D. Berghaus and D. E. Stallknecht, 2012: Persistence of low pathogenic avian influenza viruses in filtered surface water from waterfowl habitats in Georgia, USA. Journal of Wildlife Diseases,48, 999-1009.
Keeler, S. P., M. S. Dalton, A. M. Cressler, R. D. Berghaus and D. E. Stallknecht, 2014: Abiotic factors affecting the persistence of avian influenza virus in surface waters of waterfowl habitats. Applied and Enviromental Microbiology,80, 2910-2917.
Kida, H. and R. Yanagawa, 1979: Isolation and characterization of influenza a viruses from wild free-flying ducks in Hokkaido, Japan. Zentralblatt fur Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Erste Abteilung Originale. Reihe A: Medizinische Mikrobiologie und Parasitologie,244, 135-143.
Kistler, W. M., D. E. Stallknecht, T. J. Deliberto, S. Swafford, K. van Why, P. C. Wolf, J. A. Hill, D. L. Bruning, J. C. Cumbee and R. M. Mickley, 2010: Evaluation of Canada geese (Branta canadensis) as sentinels for detection of transmission of avian influenza viruses. University of Georgia.
Kleijn, D., V. J. Munster, B. S. Ebbinge, D. A. Jonkers, G. J. D. M. Müskens, Y. Van Randen and R. A. M. Fouchier, 2010: Dynamics and ecological consequences of avian influenza virus infection in greater white-fronted geese in their winter staging areas. Proceedings of the Royal Society B: Biological Sciences,277, 2041-2048.
Kocan, A. A., V. S. Hinshaw and G. A. Daubney, 1980: Influenza A Viruses Isolated from Migrating Ducks in Oklahoma Jounal of Wildlife Disease,16, 281-286.
Koehler, A. V., J. M. Pearce, P. L. Flint, J. C. Franson and H. S. Ip, 2008: Genetic evidence of intercontinental movement of avian influenza in a migratory bird: the northern pintail (Anas acuta). Molecular Ecology,17, 4754-4762.
Krauss, S., D. Walker, S. P. Pryor, L. Niles, L. Chenghong, V. S. Hinshaw and R. G. Webster, 2004: Influenza A viruses of migrating wild aquatic birds in North America. vector-Borne Zoonotic Diseases,4, 177-189.
Krauss, S., C. A. Obert, J. Franks, D. Walker, K. Jones, P. Seiler, L. Niles, S. P. Pryor, J. C. Obenauer and C. W. Naeve, 2007: Influenza in migratory birds and evidence of limited intercontinental virus exchange. PLoS Pathogens,3,
Krauss, S., D. E. Stallknecht, N. J. Negovetich, L. J. Niles, R. J. Webby and R. G. Webster, 2010: Coincident ruddy turnstone migration and horseshoe crab spawning creates an ecological ‘hot spot’for influenza viruses. Proceedings of the Royal Society B: Biological Sciences,277, 3373-3379.
Latorre-Margalef, N., M. Wille, D. Bengtsson, G. Gunnarsson, V. Grosbois, D. Hasselquist, B. Olsen, J. Elmberg and J. Waldenström, 2013: Individual variation in influenza A virus infection histories and long-term immune responses in mallards. PLoS One,8,
Latorre-Margalef, N., V. Grosbois, J. Wahlgren, V. J. Munster, C. Tolf, R. A. M. Fouchier, A. D. M. E. Osterhaus, B. Olsen and J. Waldenström, 2013: Heterosubtypic immunity to influenza A virus infections in mallards may explain existence of multiple virus subtypes. PLoS Pathogens,9,
Latorre-Margalef, N., C. Tolf, V. Grosbois, A. Avril, D. Bengtsson, M. Wille, A. D. M. E. Osterhaus, R. A. M. Fouchier, B. Olsen and J. Waldenström, 2014: Long-term variation in influenza A virus prevalence and subtype diversity in migratory mallards in northern Europe. Proceedings of the Royal Society B: Biological Sciences,281,
Lebarbenchon, C., F. Albespy, A. L. Brochet, V. Grandhomme, F. Renaud, H. Fritz, A. J. Green, F. Thomas, S. Van Der Werf and P. Aubry, 2009: Spread of avian influenza viruses by common teal (Anas crecca) in Europe. PLoS One,4,
Lebarbenchon, C., C. M. Chang, V. Grandhomme, M. Dietrich, Y. Kayser, E. Elguero, F. Renaud, F. Thomas, S. van der Werf and M. Gauthier-Clerc, 2010: Avian influenza circulation in the Camargue (south of France) during the 2006–07 season. Avian Diseases,54, 446-449.
Lebarbenchon, C., S. Sreevatsan, M. A. Ramakrishnan, R. Poulson, V. Goekjian, J. J. Di Matteo, B. Wilcox and D. E. Stallknecht, 2010: Influenza A viruses in American white pelican (Pelecanus erythrorhynchos). Jounal of Wildlife Disease,46, 1284-1289.
Lebarbenchon, C., S. Sreevatsan, T. Lefèvre, M. Yang, M. A. Ramakrishnan, J. D. Brown and D. E. Stallknecht, 2012: Reassortant influenza A viruses in wild duck populations: effects on viral shedding and persistence in water. Proceedings of the Royal Society B: Biological Sciences,279, 3967-3975.
Lebarbenchon, C., R. Poulson, K. Shannon, B. R. Wilcox, G. A. Knutsen, C. J. Cardona and D. Stallknecht, 2013: The use of feathers sampling for influenza A virus detection in wild duck populations. Avian Diseases,57, 677-680.
Lebarbenchon, C., B. R. Wilcox, R. L. Poulson, M. J. Slusher, N. B. Fedorova, D. A. Katzel, C. J. Cardona, G. A. Knutsen, D. E. Wentworth and D. E. Stallknecht, 2015: Isolation of type a influenza viruses from red-necked grebes (Podiceps grisegena). journal of Wildlife Diseases,51, 290-293.
Kwon, Y. K., S. J. Joh, M. C. Kim, Y. J. Lee, J. G. Choi, E. K. Lee, S. H. Wee, H. W. Sung, J. H. Kwon and M. I. Kang, 2005: Highly pathogenic avian influenza in magpies (Pica pica sericea) in South Korea. Journal of Wildlife Diseases,41, 618-623.
Lee, D. H., H. J. Lee, Y. N. Lee, J. K. Park, T. H. Lim, M. S. Kim, H. N. Youn, J. B. Lee, S. Y. Park and I. S. Choi, 2011: Evidence of intercontinental transfer of North American lineage avian influenza virus into Korea. infection, Genetics and Evolution,11, 232-236.
Lee, Y. J., H. M. Kang, E. K. Lee, B. M. Song, J. Jeong, Y. K. Kwon, H. R. Kim, K. J. Lee, M. S. Hong and I. Jang, 2014: Novel reassortant influenza A (H5N8) viruses, South Korea, 2014. Emerging Infectious Diseases,20,
Lewis, N. S., Z. Javakhishvili, C. A. Russell, A. Machablishvili, P. Lexmond, J. H. Verhagen, O. Vuong, T. Onashvili, M. Donduashvili and D. J. Smith, 2013: Avian influenza virus surveillance in wild birds in Georgia: 2009–2011. PLoS One,8,
Lipkind, M. A., Y. Weisman, E. Shihmanter and D. Shoham, 1979: The first isolation of animal influenza virus in Israel. Veterinary Record.
Lipkind, M., Y. Weisman, E. Shihmanter and D. Shoham, 2003: Review of the three-year studies on the ecology of avian influenza viruses in Israel. Avian Diseases,47, 69-78.
Liu, J., H. Xiao, F. Lei, Q. Zhu, K. Qin, X.-W. Zhang, X.-l. Zhang, D. Zhao, G. Wang and Y. Feng, 2005: Highly pathogenic H5N1 influenza virus infection in migratory birds. Science,309, 1206-1206.
Lu, H., A. Castro, K. Pennick, J. Liu, Q. Yang, P. Dunn, D. Weinstock and D. Henzler, 2003: Survival of avian influenza virus H7N2 in SPF chickens and their environments. Avian Diseases,47, 1015-1021.
Lvov, D., 1978: Circulation of influenza viruses in natural biocoenosis. Viruses and environment.
Mackenzie, J. S., E. C. Edwards, R. M. Holmes and V. S. Hinshaw, 1984: Isolation of ortho‐and paramyxoviruses from wild birds in Western Australia, and the characterization of novel influenza A viruses. Australian Journal of Experimental Biology and Medical Science,62, 89-99.
Makarova, N. V., N. V. Kaverin, S. Krauss, D. Senne and R. G. Webster, 1999: Transmission of Eurasian avian H2 influenza virus to shorebirds in North America. Journal of General Virology,80, 3167-3171.
Manjunath, L. H. and B. B. Mallick, 1981: Prevalence of myxo-and paramyxoviruses in birds of Himalayan region [India]. Indian Journal of Animal Sciences,51, 1139-1143.
Marchenko, V. Y., A. Y. Alekseev, K. A. Sharshov, V. N. Petrov, N. Y. Silko, I. M. Susloparov, D. Tserennorov, D. Otgonbaatar, I. A. Savchenko and A. M. Shestopalov, 2012: Ecology of influenza virus in wild bird populations in Central Asia. Avian Diseases,56, 234-237.
Mase, M., K. Tsukamoto, T. Imada, K. Imai, N. Tanimura, K. Nakamura, Y. Yamamoto, T. Hitomi, T. Kira and T. Nakai, 2005: Characterization of H5N1 influenza A viruses isolated during the 2003–2004 influenza outbreaks in Japan. Virology 332, 167-176.
Maxted, A. M., M. P. Luttrell, V. H. Goekjian, J. D. Brown, L. J. Niles, A. D. Dey, K. S. Kalasz, D. E. Swayne and D. E. Stallknecht, 2012: Avian influenza virus infection dynamics in shorebird hosts. Journal of Wildlife Diseases,48, 322-334.
Mikami, T., M. Kawamura, T. Kondo, T. Murai, M. Horiuchi, H. Kodama, H. Izawa and H. Kida, 1987: Isolation of ortho-and paramyxoviruses from migrating feral ducks in Japan. Veterinary Record,120, 417-418.
Munster, V. J., A. Wallensten, C. Baas, G. F. Rimmelzwaan, M. Schutten, B. Olsen, A. D. M. E. Osterhaus and R. A. M. Fouchier, 2005: Mallards and highly pathogenic avian influenza ancestral viruses, northern Europe. Emerging Infectious Diseases,11,
Munster, V. J., C. Baas, P. Lexmond, J. Waldenström, A. Wallensten, T. Fransson, G. F. Rimmelzwaan, W. E. P. Beyer, M. Schutten and B. Olsen, 2007: Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathogens,3,
Muzyka, D., M. Pantin-Jackwood, E. Spackman, B. Stegniy, O. Rula and P. Shutchenko, 2012: Avian influenza virus wild bird surveillance in the Azov and Black Sea regions of Ukraine (2010–2011). Avian Diseases,56, 1010-1016.
Nazir, J., R. Haumacher, A. Ike, P. Stumpf, R. Böhm and R. E. Marschang, 2010: Long-term study on tenacity of avian influenza viruses in water (distilled water, normal saline, and surface water) at different temperatures. Avian Diseases,54, 720-724.
Nemeth, N. M., N. O. Thomas, D. S. Orahood, T. D. Anderson and P. T. Oesterle, 2010: Shedding and serologic responses following primary and secondary inoculation of house sparrows (Passer domesticus) and European starlings (Sturnus vulgaris) with low-pathogenicity avian influenza virus. Avian Pathology,39, 411-418.
Nemeth, N. M., J. D. Brown, D. E. Stallknecht, E. W. Howerth, S. H. Newman and D. E. Swayne, 2013: Experimental infection of bar-headed geese (Anser indicus) and ruddy shelducks (Tadorna ferruginea) with a clade 2.3. 2 H5N1 highly pathogenic avian influenza virus. Veterinary Pathology,50, 961-970.
Nettles, V. F., J. M. Wood and R. G. Webster, 1985: Wildlife surveillance associated with an outbreak of lethal H5N2 avian influenza in domestic poultry. Avian Diseases,29, 733-741.
Newman, S. H., S. A. Iverson, J. Y. Takekawa, M. Gilbert, D. J. Prosser, N. Batbayar, T. Natsagdorj and D. C. Douglas, 2009: Migration of whooper swans and outbreaks of highly pathogenic avian influenza H5N1 virus in eastern Asia. PLoS One,4,
Nishiura, H., B. Hoye, M. Klaassen, S. Bauer and H. Heesterbeek, 2009: How to find natural reservoir hosts from endemic prevalence in a multi-host population: a case study of influenza in waterfowl. Epidemics,1, 118-128.
Ofula, V. O., A. B. Franklin, J. J. Root, H. J. Sullivan, P. Gichuki, A. Makio, W. Bulimo, B. O. Abong’o, M. Muchai and D. Schnabel, 2013: Detection of avian influenza viruses in wild waterbirds in the Rift Valley of Kenya using fecal sampling. vector-Borne Zoonotic Diseases,13, 394-400.
Olsen, B., V. J. Munster, A. Wallensten, J. Waldenström, A. D. M. E. Osterhaus and R. A. M. Fouchier, 2006: Global patterns of influenza A virus in wild birds. Science,312, 384-388.
Olson, S. H., J. Parmley, C. Soos, M. Gilbert, N. Latorre-Margalef, J. S. Hall, P. M. Hansbro, F. Leighton, V. Munster and D. Joly, 2014: Sampling strategies and biodiversity of influenza A subtypes in wild birds. PLoS One,9,
Otsuki, K., O. Takemoto, R. Fujimoto, K. Yamazaki and T. Kubota, 1984: Isolation of H5 influenza viruses from whistling swans in western Japan in November 1983. Acta Virologica,28.
Otsuki, K., O. Takemoto, R. Fujimoto, K. Yamazaki, N. Kubota, H. Hosaki, Y. Kawaoka and M. Tsubokura, 1987: Isolation of influenza A viruses from migratory waterfowls in San-in District, Western Japan, in the winter of 1982-1983. Acta Virologica,31, 439-442.
Ottis, K. and P. A. Bachmann, 1980: Occurrence of Hsw1N1 subtype influenza A viruses in wild ducks in Europe. Archives of Virology,63, 185-190.
Ottis, K. and P. A. Bachmann, 1983: Isolation and characterization of ortho‐and paramyxoviruses from feral birds in Europe. Zentralblatt für Veterinärmedizin Reihe B,30, 22-35.
Pasick, J., Y. Berhane, C. Embury-Hyatt, J. Copps, H. Kehler, K. Handel, S. Babiuk, K. Hooper-McGrevy, Y. Li and Q. M. Le, 2007: Susceptibility of Canada geese (Branta canadensis) to highly pathogenic avian influenza virus (H5N1). Emerging Infectious Diseases,13,
Pearce, J. M., A. B. Reeves, A. M. Ramey, J. W. Hupp, H. S. Ip, M. Bertram, M. J. Petrula, B. D. Scotton, K. A. Trust and B. W. Meixell, 2011: Interspecific exchange of avian influenza virus genes in Alaska: The influence of trans‐hemispheric migratory tendency and breeding ground sympatry. Molecular Ecology,20, 1015-1025.
Pereda, A. J., M. Uhart, A. A. Perez, M. E. Zaccagnini, L. La Sala, J. Decarre, A. Goijman, L. Solari, R. Suarez and M. I. Craig, 2008: Avian influenza virus isolated in wild waterfowl in Argentina: evidence of a potentially unique phylogenetic lineage in South America. Virology,378, 363-370.
Pérez-Ramírez, E., X. Gerrikagoitia, M. Barral and U. Höfle, 2010: Detection of low pathogenic avian influenza viruses in wild birds in Castilla-La Mancha (south central Spain). Veterinary Microbiology,146, 200-208.
Perkins, L. E. L. and D. E. Swayne, 2002: Pathogenicity of a Hong Kong–origin H5N1 highly pathogenic avian influenza virus for emus, geese, ducks, and pigeons. Avian Diseases,46, 53-63.
Perkins, L. E. L. and D. E. Swayne, 2002: Susceptibility of laughing gulls (Larus atricilla) to H5N1 and H5N3 highly pathogenic avian influenza viruses. Avian Diseases,46, 877-885.
Perkins, L. E. L. and D. E. Swayne, 2003: Varied pathogenicity of a Hong Kong-origin H5N1 avian influenza virus in four passerine species and budgerigars. Veterinary Pathology,40, 14-24.
Pfitzer, S., D. J. Verwoerd, G. H. Gerdes, A. E. Labuschagne, A. Erasmus, R. Manvell and C. Grund, 2000: Newcastle disease and avian influenza A virus in wild waterfowl in South Africa. Avian Diseases 655-660.
Prosser, D., L. Hungerford, R. M. Erwin, M. A. Ottinger, J. Y. Takekawa and E. Ellis, 2013: Mapping avian influenza transmission risk at the interface of domestic poultry and wild birds. Frontiers in Public Health,1,
Ramey, A. M., J. M. Pearce, C. R. Ely, L. M. S. Guy, D. B. Irons, D. V. Derksen and H. S. Ip, 2010: Transmission and reassortment of avian influenza viruses at the Asian–North American interface. Virology,406, 352-359.
Ramey, A. M., J. M. Pearce, A. B. Reeves, J. C. Franson, M. R. Petersen and H. S. Ip, 2011: Evidence for limited exchange of avian influenza viruses between seaducks and dabbling ducks at Alaska Peninsula coastal lagoons. Archives of Virology,156, 1813-1821.
Ramey, A. M., E. Spackman, J.-Y. Yeh, G. Fujita, K. Konishi, K. Uchida, J. A. Reed, B. R. Wilcox, J. D. Brown and D. E. Stallknecht, 2013: Antibodies to H5 subtype avian influenza virus and Japanese encephalitis virus in northern pintails (Anas acuta) sampled in Japan. Japanese Journal of Veterinary Research,61, 117-123.
Ramey, A., N. Latorre-Margalef, R. Poulson, A. González-Reiche, D. Perez, D. Stallknecht and J. Brown, 2014: Genomic, epidemiologic, and pathogenic characteristics of H14 subtype influenza A viruses in New World waterfowl. PLoS One,9,
Ramey, A. M., P. Walther, P. Link, R. L. Poulson, B. R. Wilcox, G. Newsome, E. Spackman, J. D. Brown and D. E. Stallknecht, 2016: Optimizing Surveillance for South American Origin Influenza A Viruses Along the United States Gulf Coast Through Genomic Characterization of Isolates from Blue‐winged Teal (Anas discors). Transboundary and Emerging Diseases,63, 194-202.
Ramey, A., R. Poulson, A. González-Reiche, B. Wilcox, P. Walther, P. Link, D. Carter, G. Newsome, M. Müller and R. Berghaus, 2014: Evidence for seasonal patterns in the relative abundance of avian influenza virus subtypes in North American waterfowl. Journal of Wildlife Diseases,50, 916-922.
Reeves, A. B., J. M. Pearce, A. M. Ramey, B. W. Meixell and J. A. Runstadler, 2011: Interspecies transmission and limited persistence of low pathogenic avian influenza genomes among Alaska dabbling ducks. Infection, Genetics and Evolution,11, 2004-2010.
Rohani, P., R. Breban, D. Stallknecht and J. M. Drake, 2009: Environmental transmission of avian influenza viruses and its implications for disease control. Proceedings of the National Academy of Sciences of the United States of America,106, 10365-10369.
Romvary, J. and J. Tanyi, 1975: Occurrence of Hong Kong influenza A (H3N2) virus infection in the Budapest Zoo. Acta Veterinaria Academiae Scientiarum Hungariae,25, 251-254.
Romváry, J., J. Meszaros, J. Tanyi, J. Rosza and L. Fabian, 1976: Influenza infectedness of captured and shot wild birds on north-eastern and south-eastern parts of Hungary. Acta Veterinaria Academiae Scientiarum Hungariae,26, 363-368.
Romvary, J., J. Meszaros, J. Tanyi, J. Rosza and L. Fabian, 1976: Spreading of virus infection among wild birds and monkeys during the influenza epidemic caused by the Victoria (3) 75 variant of a (H3N2) virus. Acta Veterinaria Academiae Scientiarum Hungariae,26, 369-376.
Rosenberger, J., W. Krauss and R. D. Slemons, 1974: Isolation of Newcastle disease and type-A influenza viruses from migratory waterfowl in the Atlantic flyway. Avian Diseases,18, 610-613.
Roslaia, I. G., G. E. Roslaiakov and D. K. Lvov, 1975: Isolation of influenza A viruses and detection of antibodies in common herons (Ardea cinera) nesting in the lower Amur. Ekol Virusov,3, 138-142.
Roslaya, I., G. Roslyakov, D. Lvov, V. Isacenko, L. Y. Zakstelskaya, L. Ya and I. Trop, 1974: Circulation of arbo-and myxoviruses in populations of waterfowl and shore birds of the lower Amur [in Russian]. Ivanovsky Institute of Virology,2, 148-156.
Runstadler, J., N. Hill, I. T. Hussein, W. Puryear and M. Keogh, 2013: Connecting the study of wild influenza with the potential for pandemic disease. Infection, Genetics and Evolution,17, 162-187.
Sabirovic, M., J. Wilesmith, S. Hall, N. Coulson and F. Landeg, 2006: Outbreaks of HPAI H5N1 virus in Europe during 2005/2006-An overview and commentary. International Animal Health Division: London, UK.
Saito, T., T. Horimoto, Y. Kawaoka, D. A. Senne and R. G. Webster, 1994: Emergence of a potentially pathogenic H5N2 influenza virus in chickens. Virology,201, 277-284.
Sandhu, T. and V. Hinshaw, 2003: Influenza A virus infection of domestic ducks. Proceedings of the First International Conference on Avian Influenza, A. Bankowski, ed. Carter Composition Corp,; Richmond, VA, 93-99
Sazonov, A. A., D. K. Lvov, R. G. Webster, T. V. Sokolova, N. A. Braude and N. V. Portyanko, 1977: Isolation of an influenza virus, similar to A/Port Chalmers/1/73 (H3N2) from a common murre at Sakhalin Island in USSR (Strain A/CommonMurre/Sakhalin/1/74). Archives of Virology,53, 1-7.
Schäffr, J. R., Y. Kawaoka, W. J. Bean, J. Süss, D. Senne and R. G. Webster, 1993: Origin of the pandemic 1957 H2 influenza A virus and the persistence of its possible progenitors in the avian reservoir. Virology,194, 781-788.
Senne, D. A., J. E. Pearson, L. D. Miller and G. A. Gustafson, 1983: Virus isolations from pet birds submitted for importation into the United States. Avian Diseases,27, 731-744.
Sharp, G., Y. Kawaoka, S. Wright, B. Turner, V. Hinshaw and R. Webster, 1993: Wild ducks are the reservoir for only a limited number of influenza A subtypes. Epidemiology and Infection,110, 161-176.
Sinnecker, R., H. Sinnecker, E. Zilske and D. Köhler, 1983: Surveillance of pelagic birds for influenza A viruses. Acta Virologica,27, 75-79.
Slemons, R. D. and B. Easterday, 1972: Host response differences among 5 avian species to an influenzavirus—A/turkey/Ontario/7732/66 (Hav5N?). Bulletin of the World Health Organization,47,
Slemons, R. and B. C. Easterday, 1975. The natural history of type-A influenza viruses and wild waterfowl. In: Third International Wildlife Disease Conference, L. A. Page, ed. Plenum Press; New York, 215-224.
Slemons, R. D., D. C. Johnson, J. S. Osborn and F. Hayes, 1974: Type-A influenza viruses isolated from wild free-flying ducks in California. Avian Diseases, 119-124.
Slemons, R., M. Shieldcastle, L. Heyman, K. Bednarik and D. Senne, 1991: Type A influenza viruses in waterfowl in Ohio and implications for domestic turkeys. Avian Diseases,18, 165-173.
Slusher, M. J., B. R. Wilcox, M. P. Lutrell, R. L. Poulson, J. D. Brown, M. J. Yabsley and D. E. Stallknecht, 2014: Are passerine birds reservoirs for influenza A viruses? Journal of Wildlife Diseases,50, 792-809.
Smitka, C., 1981: Ortho-and paramyxoviruses in the migratory waterfowl of Michigan. Journal of Wildlife Diseases,17, 147-151.
Spackman, E., D. E. Stallknecht, R. D. Slemons, K. Winker, D. L. Suarez, M. Scott and D. E. Swayne, 2005: Phylogenetic analyses of type A influenza genes in natural reservoir species in North America reveals genetic variation. Virus Research,114, 89-100.
Spackman, E., K. G. McCracken, K. Winker and D. E. Swayne, 2007: An avian influenza virus from waterfowl in South America contains genes from North American avian and equine lineages. Avian Diseases,51, 273-274.
Spackman, E., D. E. Swayne, M. Gilbert, D. O. Joly, W. B. Karesh, D. L. Suarez, R. Sodnomdarjaa, P. Dulam and C. J. V. j. Cardona, 2009: Characterization of low pathogenicity avian influenza viruses isolated from wild birds in Mongolia 2005 through 2007. 6, 1-8.
Squires, R. B., J. Noronha, V. Hunt, A. García‐Sastre, C. Macken, N. Baumgarth, D. Suarez, B. E. Pickett, Y. Zhang and C. N. Larsen, 2012: Influenza research database: an integrated bioinformatics resource for influenza research and surveillance. Influenza and Other Respiratory Viruses,6, 404-416.
Stallknecht, D. E. and S. Shane, 1988: Host range of avian influenza virus in free-living birds. Veterinary Research Communications,12, 125-141.
Stallknecht, D. E., S. Shane, P. Zwank, D. Senne and M. Kearney, 1990: Avian influenza viruses from migratory and resident ducks of coastal Louisiana. Avian Diseases,34, 398-405.
Stallknecht, D., S. Shane, M. Kearney and P. Zwank, 1990: Persistence of avian influenza viruses in water. Avian Diseases, 406-411.
Stallknecht, D., M. Kearney, S. Shane and P. Zwank, 1990: Effects of pH, temperature, and salinity on persistence of avian influenza viruses in water. Avian Diseases, 412-418.
Stallknecht, D. E., 2003: Ecology and epidemiology of avian influenza viruses in wild bird populations: waterfowl, shorebirds, pelicans, cormorants, etc. Avian Diseases,47, 61-69. (Special Issue: Proceedings of Fourth International Symposium on Avian Influenza, 1997):61-69
Stallknecht, D. E., J. D. Brown and D. Swayne, 2008: Ecology of avian influenza in wild birds. In: D. E. Swayne (ed.), Avian Influenza. Blackwell Publishing, Ames, IA.
Stallknecht, D. E., V. H. Goekjian, B. R. Wilcox, R. L. Poulson and J. D. Brown, 2010: Avian influenza virus in aquatic habitats: what do we need to learn? Avian Diseases,54, 461-465.
Stallknecht, D., M. Luttrell, R. Poulson, V. Goekjian, L. Niles, A. Dey, S. Krauss and R. Webster, 2012: Detection of avian influenza viruses from shorebirds: evaluation of surveillance and testing approaches. Journal of Wildlife Diseases,48, 382-393.
Stanislawek, W., C. Wilks, J. Meers, G. Horner, D. Alexander, R. Manvell, J. Kattenbelt and A. Gould, 2002: Avian paramyxoviruses and influenza viruses isolated from mallard ducks (Anas platyrhynchos) in New Zealand. Archives of Virology,147, 1287-1302.
StüNzner, D., W. Thiel, F. Pötsch and W. Sixl, 1980: Isolation of influenza viruses from exotic and central European birds. Zentralblatt für Bakteriologie. 1. Abt. Originale A, Medizinische Mikrobiologie, Infektionskrankheiten und Parasitologie,247, 8-17.
Sturm-Ramirez, K. M., T. Ellis, B. Bousfield, L. Bissett, K. Dyrting, J. E. Rehg, L. Poon, Y. Guan, M. Peiris and R. G. Webster, 2004: Reemerging H5N1 influenza viruses in Hong Kong in 2002 are highly pathogenic to ducks. Journal of Virology,78, 4892-4901.
Süss, J., J. Schäfer, H. Sinnecker and R. Webster, 1994: Influenza virus subtypes in aquatic birds of eastern Germany. Archives of Virology,135, 101-114.
Takekawa, J. Y., D. J. Prosser, B. M. Collins, D. C. Douglas, W. M. Perry, B. Yan, L. Ze, Y. Hou, F. Lei and T. Li, 2013: Movements of wild ruddy shelducks in the Central Asian Flyway and their spatial relationship to outbreaks of highly pathogenic avian influenza H5N1. Viruses,5, 2129-2152.
Thorsen, J., I. K. Barker and V. S. Hinshaw, 1980: Influenza viruses isolated from waterfowl in southern Ontario, 1976–1978. Canadian Journal of Microbiology,26, 1511-1514.
Tolf, C., D. Bengtsson, D. Rodrigues, N. Latorre-Margalef, M. Wille, M. E. Figueiredo, M. Jankowska-Hjortaas, A. Germundsson, P.-Y. Duby and C. Lebarbenchon, 2012: Birds and viruses at a crossroad-surveillance of influenza A virus in Portuguese waterfowl. PLoS One,7,
Tolf, C., N. Latorre-Margalef, M. Wille, D. Bengtsson, G. Gunnarsson, V. Grosbois, D. Hasselquist, B. Olsen, J. Elmberg and J. Waldenström, 2013: Individual variation in influenza A virus infection histories and long-term immune responses in mallards. PLoS One,8,
Toma, B., J.-J. C. Benet, B. C. Duford, M. C. Eloit, W. C. Marsh, P. C. Michel, F. C. Moutou, M. C. Sanaa and J.-P. Vaillancourt, 1999: Dictionary of veterinary epidemiology. Iowa State University Press.
Tong, S., Y. Li, P. Rivailler, C. Conrardy, D. A. A. Castillo, L.-M. Chen, S. Recuenco, J. A. Ellison, C. T. Davis and I. A. York, 2012: A distinct lineage of influenza A virus from bats. Proceedings of the National Academy of Sciences,109, 4269-4274.
Tong, S., X. Zhu, Y. Li, M. Shi, J. Zhang, M. Bourgeois, H. Yang, X. Chen, S. Recuenco and J. Gomez, 2013: New world bats harbor diverse influenza A viruses. PLoS Pathogens,9,
Toennessen, R., A. Germundsson, C. M. Jonassen, I. Haugen, K. Berg, R. T. Barrett and E. Rimstad, 2011: Virological and serological surveillance for type A influenza in the black-legged kittiwake (Rissa tridactyla). Virology Journal,8, 1-8.
Tønnessen, R., A. B. Kristoffersen, C. M. Jonassen, M. J. Hjortaas, E. F. Hansen, E. Rimstad and A. G. Hauge, 2013: Molecular and epidemiological characterization of avian influenza viruses from gulls and dabbling ducks in Norway. Virology Journal,10, 1-10.
Tracey, J. P., R. Woods, D. Roshier, P. West and G. R. Saunders, 2004: The role of wild birds in the transmission of avian influenza for Australia: an ecological perspective. Emu,104, 109-124.
Tsubokura, M., K. Otsuki, Y. Kawaoka and R. Yanagawa, 1981: Isolation of influenza A viruses from migratory waterfowls in San-in District, Western Japan in 1979-1980. Zentralblatt fur Bakteriologie, Mikrobiologie und Hygiene. 1. Abt. Originale B, Hygiene,173, 494-500.
Tsubokura, M., K. Otsuki, H. Yamamoto, Y. Kawaoka and K. Nerome, 1981: Isolation of an Hswl Nav4 influenza virus from a tufted duck (Aythya fuligula) in Japan. Microbiology and Immunology,25, 819-825.
Turek, R., B. Tůmová, V. Mucha and A. Stumpa, 1983: Type A influenza virus strains isolated from free living ducks in Czechoslovakia during 1978-1981. Acta Virologica,27, 523-527.
Van Dijk, J. G., B. J. Hoye, J. H. Verhagen, B. A. Nolet, R. A. Fouchier and M. Klaassen, 2014: Juveniles and migrants as drivers for seasonal epizootics of avian influenza virus. Journal of Animal Ecology,83, 266-275.
Velarde, R., S. E. Calvin, D. Ojkic, I. K. Barker and É. Nagy, 2010: Avian influenza virus H13 circulating in ring-billed gulls (Larus delawarensis) in southern Ontario, Canada. Avian Diseases,54, 411-419.
Verhagen, J. H., F. Majoor, P. Lexmond, O. Vuong, G. Kasemir, D. Lutterop, A. D. Osterhaus, R. A. Fouchier and T. Kuiken, 2014: Epidemiology of influenza A virus among black-headed gulls, the Netherlands, 2006–2010. Emerging Infectious Diseases,20,
Verhagen, J. H., J. G. van Dijk, O. Vuong, T. Bestebroer, P. Lexmond, M. Klaassen and R. A. Fouchier, 2014: Migratory birds reinforce local circulation of avian influenza viruses. PLoS One,9,
Vijaykrishna, D., Y.-M. Deng, Y. C. Su, M. Fourment, P. Iannello, G. G. Arzey, P. M. Hansbro, K. E. Arzey, P. D. Kirkland and S. Warner, 2013: The recent establishment of North American H10 lineage influenza viruses in Australian wild waterfowl and the evolution of Australian avian influenza viruses. Journal of virology,87, 10182-10189.
Wallensten, A., V. J. Munster, M. Karlsson, Å. Lundkvist, M. Brytting, M. Stervander, A. D. Osterhaus, R. A. Fouchier and B. Olsen, 2006: High prevalence of influenza A virus in ducks caught during spring migration through Sweden. Vaccine,24, 6734-6735.
Wallensten, A., V. J. Munster, N. Latorre-Margalef, M. Brytting, J. Elmberg, R. A. Fouchier, T. Fransson, P. D. Haemig, M. Karlsson and Å. Lundkvist, 2007: Surveillance of influenza virus A in migratory waterfowl in Northern Europe. Emerging Infectious Diseases,13,
Watanabe, T., M. Kiso, S. Fukuyama, N. Nakajima, M. Imai, S. Yamada, S. Murakami, S. Yamayoshi, K. Iwatsuki-Horimoto and Y. Sakoda, 2013: Characterization of H7N9 influenza A viruses isolated from humans. Nature,501, 551-555.
Webster, R., M. Morita, C. Pridgen and B. Tumova, 1976: Ortho-and paramyxoviruses from migrating feral ducks: characterization of a new group of influenza A viruses. Journal of General Virology,32, 217-225.
Webster, R. G., M. Yakhno, V. S. Hinshaw, W. J. Bean and K. C. Murti, 1978: Intestinal influenza: replication and characterization of influenza viruses in ducks. Virology Journal,84, 268-278.
Webster, R., V. Hinshaw, W. Bean, K. Van Wyke, J. Geraci, D. S. Aubin and G. Petursson, 1981: Characterization of an influenza A virus from seals. Virology,113, 712-724.
Wilcox, B. R., G. A. Knutsen, J. Berdeen, V. Goekjian, R. Poulson, S. Goyal, S. Sreevatsan, C. Cardona, R. D. Berghaus and D. E. Swayne, 2011: Influenza-A viruses in ducks in northwestern Minnesota: fine scale spatial and temporal variation in prevalence and subtype diversity. PloS one,6,
Wille, M., G. J. Robertson, H. Whitney, D. Ojkic and A. S. Lang, 2011: Reassortment of American and Eurasian genes in an influenza A virus isolated from a great black-backed gull (Larus marinus), a species demonstrated to move between these regions. Archives of Virology,156, 107-115.
Wille, M., C. Tolf, A. Avril, N. Latorre-Margalef, S. Wallerström, B. Olsen and J. Waldenström, 2013: Frequency and patterns of reassortment in natural influenza A virus infection in a reservoir host. Virology,443, 150-160.
Wood, J., R. Webster and V. Nettles, 1985: Host range of A/Chicken/Pennsylvania/83 (H5N2) influenza virus. Avian Diseases, 198-207.
Yamane, N., T. Odagiri, J. Arikawa and N. Ishida, 1979: Isolation and characterization of Influenza A viruses from wild ducks in northern Japan: appearance of Hsw1 antigens in the Japanese duck population. Acta Virologica,23, 375-384.
Yamane, N., T. Odagiri, J. Arikawa, M. Morita, N. Sukeno and N. Ishida, 1978: Isolation of orthomyxoviruses from migrating and domestic ducks in northern Japan in 1976-1977. Japanese Journal of Medical Science and Biology,31, 407-415.
Zakstelskaya, L. Y., M. Yakhno, V. Isachenko, S. Klimenko, E. Molibog, V. Andreev, D. Lvov and S. Yamnikova, 1974: Isolation and peculiarities of influenza virus (tern)(Turkmenistan 18) 73. Virologiia Virusov,2, 93-98.
DISCLAIMER
This article was translated to enable Bahasa Indonesia readers to better understand the topic explained inside the article in their native language. The translator made no profit in translating the article.